 Le microbiologiste Carl R. Woese et des scientifiques de l’Université de l’Illinois annoncent l’identification des méthanogènes, une forme de vie microbienne (Archaea) datant d’environ 3,5 milliards d’années
Le microbiologiste Carl R. Woese et des scientifiques de l’Université de l’Illinois annoncent l’identification des méthanogènes, une forme de vie microbienne (Archaea) datant d’environ 3,5 milliards d’années 
 Les bactéries méthanogènes sont uniques parmi les procaryotes car elles produisent un hydrocarbure, le méthane, comme produit majeur du métabolisme anaérobie. Cette propriété physiologique a été proposée en 1956 par HA Barker comme caractéristique taxonomique principale d’un groupe morphologiquement diversifié de bactéries qu’il a appelé les Methanobacteriaceae. La taxonomie de cette famille physiologique a été obscurcie par la difficulté d’obtenir des membres en culture pure. Par conséquent, diverses espèces ont été nommées en fonction des types de substrats convertis en méthane par des cultures « purifiées » (c’est-à-dire enrichies) contenant un type morphologique prédominant suspecté de méthanogenèse. Seulement trois espèces, Methanobacterium formicicum, Methano-coccus vannielii et Methanosarcina barkeri, ont été isolés en culture axénique au moment de l’examen de ces organismes par Barker
Les bactéries méthanogènes sont uniques parmi les procaryotes car elles produisent un hydrocarbure, le méthane, comme produit majeur du métabolisme anaérobie. Cette propriété physiologique a été proposée en 1956 par HA Barker comme caractéristique taxonomique principale d’un groupe morphologiquement diversifié de bactéries qu’il a appelé les Methanobacteriaceae. La taxonomie de cette famille physiologique a été obscurcie par la difficulté d’obtenir des membres en culture pure. Par conséquent, diverses espèces ont été nommées en fonction des types de substrats convertis en méthane par des cultures « purifiées » (c’est-à-dire enrichies) contenant un type morphologique prédominant suspecté de méthanogenèse. Seulement trois espèces, Methanobacterium formicicum, Methano-coccus vannielii et Methanosarcina barkeri, ont été isolés en culture axénique au moment de l’examen de ces organismes par Barker  Les bactéries méthanogènes forment un groupe important et diversifié qui est uni par trois caractéristiques :
Les bactéries méthanogènes forment un groupe important et diversifié qui est uni par trois caractéristiques :
(1) Elles forment de grandes quantités de méthane comme principal produit de leur métabolisme énergétique. (2) Ce sont des anaérobies stricts.
(3) Ils sont membres du domaine Archaea, ou archaebactéries, et ne sont apparentés que de loin aux bactéries ou eubactéries classiques plus familières.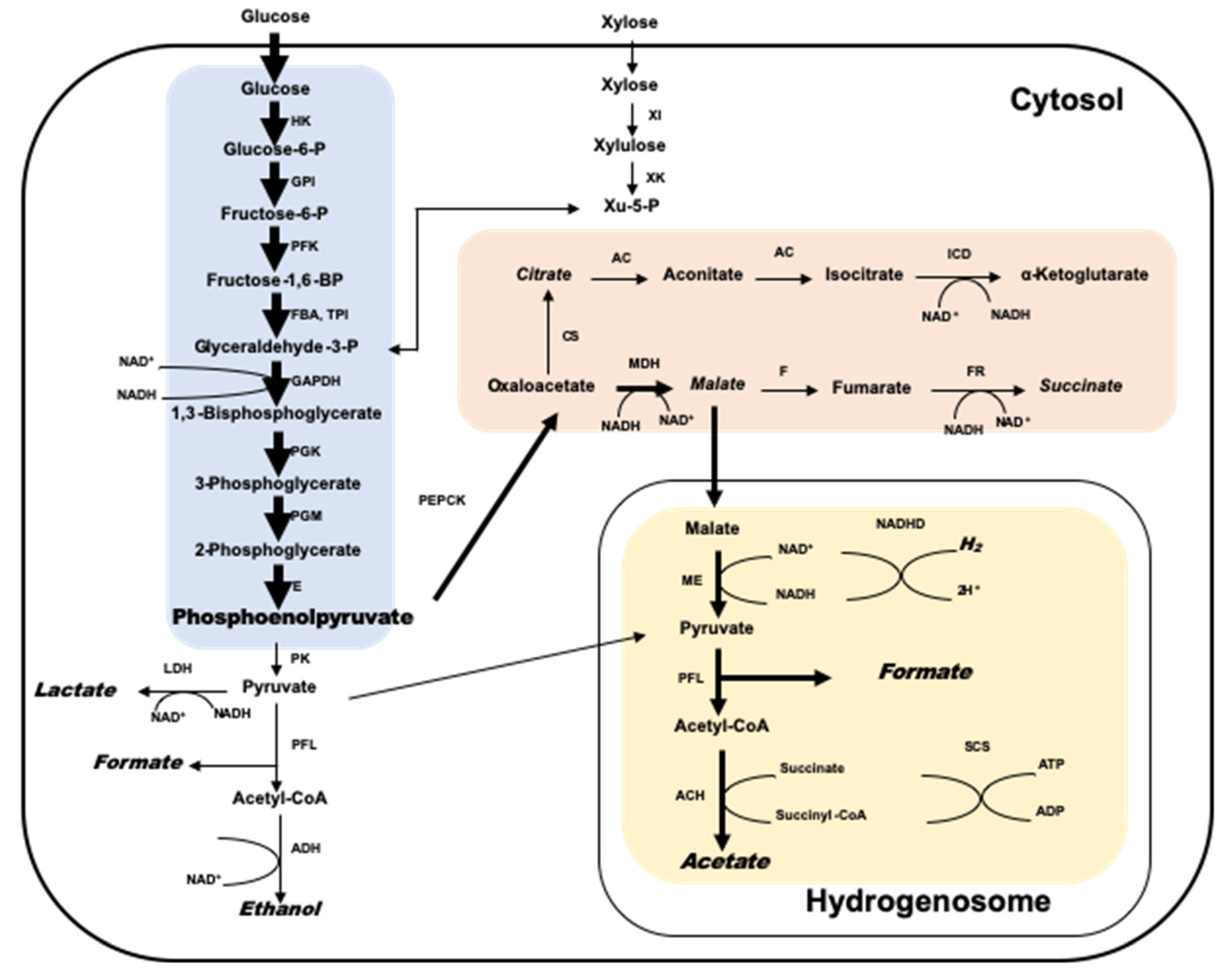 Comme les eubactéries photosynthétiques, les bactéries méthanogènes sont liées entre elles principalement par leur mode de métabolisme énergétique mais sont très diverses quant à leurs autres propriétés. Les bactéries méthanogènes tirent leur énergie de croissance de la conversion d’un nombre limité de substrats en gaz méthane. Les principaux substrats sont H2 + CO2, formiate et acétate. De plus, certains autres composés C-1 tels que le méthanol, la triméthylamine et le sulfure de diméthyle et certains alcools tels que l’isopropanol, l’isobutanol, le cyclopentanol et l’éthanol sont des substrats pour certains méthanogènes. Tous ces substrats sont transformés…
Comme les eubactéries photosynthétiques, les bactéries méthanogènes sont liées entre elles principalement par leur mode de métabolisme énergétique mais sont très diverses quant à leurs autres propriétés. Les bactéries méthanogènes tirent leur énergie de croissance de la conversion d’un nombre limité de substrats en gaz méthane. Les principaux substrats sont H2 + CO2, formiate et acétate. De plus, certains autres composés C-1 tels que le méthanol, la triméthylamine et le sulfure de diméthyle et certains alcools tels que l’isopropanol, l’isobutanol, le cyclopentanol et l’éthanol sont des substrats pour certains méthanogènes. Tous ces substrats sont transformés… CO dans la méthanogenèse
CO dans la méthanogenèse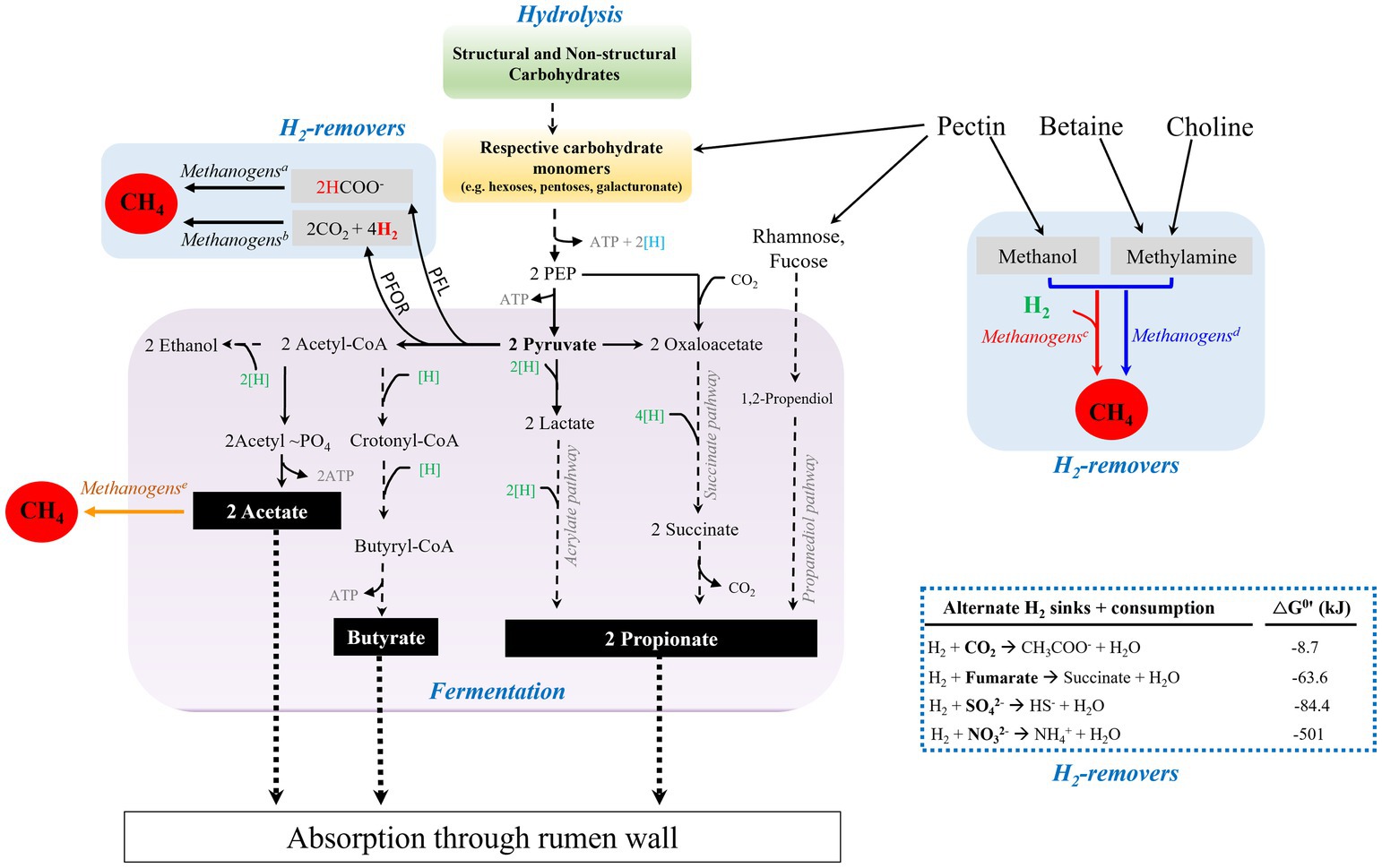
Bien que le CO soit présent dans les environnements méthanogènes, la compréhension du métabolisme du CO par les méthanogènes est à la traîne par rapport aux autres substrats méthanogènes et aux enquêtes sur le métabolisme du CO chez les espèces non méthanogènes. Cette revue présente des études sur le métabolisme du CO par les méthanogènes de 1931 à nos jours. Les voies du métabolisme du CO des espèces d’eau douce par rapport aux espèces marines sont contrastées et les implications écologiques discutées. La biochimie et le rôle de la CO déshydrogénase/acétyl-CoA synthase dans la voie de conversion de l’acétate en méthane et la biosynthèse du carbone cellulaire sont présentés. Enfin, une proposition sur le rôle du CO et des formes primitives de la CO déshydrogénase/acétyl-CoA synthase dans l’origine et l’évolution précoce de la vie est discutée. Introduction
Introduction
La décomposition de la matière organique complexe dans divers environnements anaérobies est un maillon essentiel du cycle global du carbone (Fig. 1) produisant environ 1 milliard de tonnes de méthane chaque année. Dans le cycle, le CO2 est fixé dans la matière organique complexe par la photosynthèse qui est décomposée principalement par des micro-organismes aérobies nécessitant de l’O2 dans des habitats oxygénés avec libération de CO2 dans l’atmosphère. Cependant, une partie de la matière organique est également déposée dans divers habitats exempts d’O2 où les chaînes alimentaires microbiennes anaérobies décomposent la matière organique en CH4 et CO2 dans un processus appelé biométhanisation. Une partie du CH4 est reconverti en CO2 par les méthylotrophes anaérobies et le reste s’échappe dans les zones aérobies où il est oxydé en CO2 par les méthylotrophes nécessitant de l’O2, complétant ainsi le cycle global du carbone.
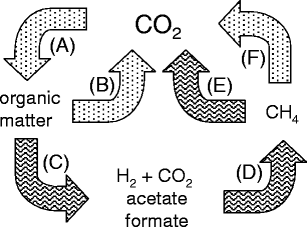
Fig. 1 – Le cycle mondial du carbone. a Fixation du CO2 dans la matière organique, b décomposition aérobie de la matière organique en CO2, c décomposition anaérobie de la matière organique en produits finis de fermentation, d conversion anaérobie des produits finis de fermentation en CH4, e oxydation anaérobie du CH4 en CO2, f oxydation aérobie du CH 4 en CO2 La biométhanisation de la matière organique se produit dans divers habitats tels que les sédiments d’eau douce, les rizières, les digesteurs d’eaux usées, le rumen, le tractus intestinal inférieur des animaux monogastriques, les décharges, les cheminées hydrothermales, les sédiments marins côtiers et le sous-sol (Liu et Whitman 2008). Un minimum de trois groupes métaboliques d’anaérobies en interaction forment un consortium convertissant la matière organique complexe en CO2 et CH4 (Fig. 2). Les anaérobies fermentaires du groupe I décomposent la matière organique complexe en acétate, en acides gras volatils plus élevés, en H2 et en CO2. Les anaérobies acétogènes du groupe II producteurs de H2 décomposent les acides gras les plus volatils en acétate, H2 et C02.
La biométhanisation de la matière organique se produit dans divers habitats tels que les sédiments d’eau douce, les rizières, les digesteurs d’eaux usées, le rumen, le tractus intestinal inférieur des animaux monogastriques, les décharges, les cheminées hydrothermales, les sédiments marins côtiers et le sous-sol (Liu et Whitman 2008). Un minimum de trois groupes métaboliques d’anaérobies en interaction forment un consortium convertissant la matière organique complexe en CO2 et CH4 (Fig. 2). Les anaérobies fermentaires du groupe I décomposent la matière organique complexe en acétate, en acides gras volatils plus élevés, en H2 et en CO2. Les anaérobies acétogènes du groupe II producteurs de H2 décomposent les acides gras les plus volatils en acétate, H2 et C02.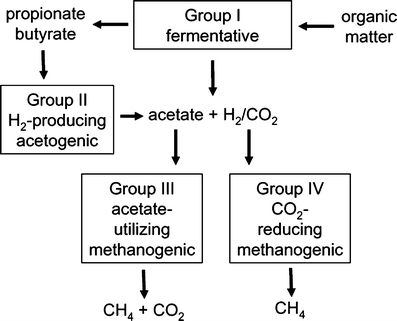 Les méthanogènes des groupes III et IV convertissent les produits métaboliques des deux premiers groupes en CH4 par deux voies principales. Au moins les deux tiers du CH4 produit dans la nature proviennent de la conversion du groupe méthyle de l’acétate par les méthanogènes du groupe III. Les méthanogènes du groupe IV en produisent environ un tiers en réduisant le CO2 avec des électrons fournis par l’oxydation de H2 ou de formiate. Ainsi, les méthanogènes dépendent des deux premiers groupes pour fournir des substrats à leur croissance.
Les méthanogènes des groupes III et IV convertissent les produits métaboliques des deux premiers groupes en CH4 par deux voies principales. Au moins les deux tiers du CH4 produit dans la nature proviennent de la conversion du groupe méthyle de l’acétate par les méthanogènes du groupe III. Les méthanogènes du groupe IV en produisent environ un tiers en réduisant le CO2 avec des électrons fournis par l’oxydation de H2 ou de formiate. Ainsi, les méthanogènes dépendent des deux premiers groupes pour fournir des substrats à leur croissance. 
 Le CO comme source d’énergie
Le CO comme source d’énergie

Le premier rapport enregistré laissant présager la capacité des méthanogènes à métaboliser le CO est apparu en 1931 lorsqu’il a été démontré que les boues d’épuration convertissent le CO en CO2 et CH4 (Fischer et al. 1931). Cependant, la conversion directe du CO par les méthanogènes (Eq.1) n’a pas pu être conclue car le H2 et l’acétate sont apparus et ont disparu conformément aux non-méthanogènes en tant que principaux anaérobies métabolisant le CO en substrats méthanogènes En 1933, il a été rapporté qu’une culture pure était capable de convertir le CO en CH4 en convertissant d’abord le CO en H2 qui, 14 ans plus tard en 1947, a été confirmé avec des cellules au repos suspensions d’une culture pure. Un autre décalage dans le temps s’était écoulé avant que les études sur le métabolisme du CO par les méthanogènes ne reprennent en 1977 et 1984 lorsqu’il a été démontré que plusieurs espèces éliminent le CO de la phase gazeuse lors de la croissance avec CO2 plus H2 et, pour la première fois, que deux méthanogènes d’eau douce sont capables de se développer avec le CO comme seule source d’énergie, bien que dans des conditions incapables de soutenir la prolifération dans l’environnement natif. Ce n’est que récemment qu’il a été démontré qu’une espèce marine se développait avec le CO dans des conditions propices à la prolifération dans l’environnement natif via une nouvelle voie de méthanogenèse compatible avec la croissance dépendante du CO dans l’environnement natif.
En 1933, il a été rapporté qu’une culture pure était capable de convertir le CO en CH4 en convertissant d’abord le CO en H2 qui, 14 ans plus tard en 1947, a été confirmé avec des cellules au repos suspensions d’une culture pure. Un autre décalage dans le temps s’était écoulé avant que les études sur le métabolisme du CO par les méthanogènes ne reprennent en 1977 et 1984 lorsqu’il a été démontré que plusieurs espèces éliminent le CO de la phase gazeuse lors de la croissance avec CO2 plus H2 et, pour la première fois, que deux méthanogènes d’eau douce sont capables de se développer avec le CO comme seule source d’énergie, bien que dans des conditions incapables de soutenir la prolifération dans l’environnement natif. Ce n’est que récemment qu’il a été démontré qu’une espèce marine se développait avec le CO dans des conditions propices à la prolifération dans l’environnement natif via une nouvelle voie de méthanogenèse compatible avec la croissance dépendante du CO dans l’environnement natif. Archaebactéries : le troisième domaine de la vie manqué par les biologistes pendant des décennies
Archaebactéries : le troisième domaine de la vie manqué par les biologistes pendant des décennies

Ces bactéries inhabituelles ne sont généalogiquement ni procaryotes ni eucaryotes. Cette découverte signifie qu’il n’y a pas deux lignées de descendance de la vie mais trois : les archaebactéries, les vraies bactéries et les eucaryotes Les premiers philosophes de la nature soutenaient que la vie sur terre est fondamentalement dichotomique : tous les êtres vivants sont soit des animaux, soit des plantes. Lorsque les micro-organismes ont été découverts, ils ont été divisés de la même manière. Les grands et mobiles étaient considérés comme des animaux et ceux qui semblaient ne pas bouger, y compris les bactéries, étaient considérés comme des plantes. Au fur et à mesure que la compréhension du monde microscopique progressait, il est devenu évident qu’une simple classification en deux volets ne suffirait pas, et ainsi des catégories supplémentaires ont été introduites : champignons, protozoaires et bactéries.
Les premiers philosophes de la nature soutenaient que la vie sur terre est fondamentalement dichotomique : tous les êtres vivants sont soit des animaux, soit des plantes. Lorsque les micro-organismes ont été découverts, ils ont été divisés de la même manière. Les grands et mobiles étaient considérés comme des animaux et ceux qui semblaient ne pas bouger, y compris les bactéries, étaient considérés comme des plantes. Au fur et à mesure que la compréhension du monde microscopique progressait, il est devenu évident qu’une simple classification en deux volets ne suffirait pas, et ainsi des catégories supplémentaires ont été introduites : champignons, protozoaires et bactéries.  En fin de compte, cependant, une nouvelle simplification s’est imposée. Il semblait que la vie pouvait être dichotomique après tout, mais à un niveau plus profond, à savoir dans la structure de la cellule vivante. Toutes les cellules semblaient appartenir à l’un ou l’autre de deux groupes : les eucaryotes, qui sont des cellules à noyau bien formé, et les procaryotes, qui n’ont pas un tel noyau. Les plantes et les animaux multicellulaires sont eucaryotes, tout comme de nombreux organismes unicellulaires. Les seuls procaryotes sont les bactéries (dont les cyanobactéries, autrefois appelées algues bleues).
En fin de compte, cependant, une nouvelle simplification s’est imposée. Il semblait que la vie pouvait être dichotomique après tout, mais à un niveau plus profond, à savoir dans la structure de la cellule vivante. Toutes les cellules semblaient appartenir à l’un ou l’autre de deux groupes : les eucaryotes, qui sont des cellules à noyau bien formé, et les procaryotes, qui n’ont pas un tel noyau. Les plantes et les animaux multicellulaires sont eucaryotes, tout comme de nombreux organismes unicellulaires. Les seuls procaryotes sont les bactéries (dont les cyanobactéries, autrefois appelées algues bleues). Ces dernières années, mes collègues et moi avons été amenés à proposer une révision fondamentale de ce tableau. Parmi les bactéries, nous avons trouvé un groupe d’organismes qui ne semblent appartenir à aucune des catégories de base. Les organismes que nous avons étudiés sont procaryotes en ce sens qu’ils n’ont pas de noyau et qu’ils ressemblent beaucoup à des bactéries ordinaires. Cependant, dans leur biochimie et dans la structure de certaines grosses molécules, ils sont aussi différents des autres procaryotes que des eucaryotes. Phylogénétiquement, ils ne sont ni procaryotes ni eucaryotes. Ils constituent un nouveau « royaume primaire », avec un statut complètement différent dans l’histoire et l’ordre naturel de la vie.
Ces dernières années, mes collègues et moi avons été amenés à proposer une révision fondamentale de ce tableau. Parmi les bactéries, nous avons trouvé un groupe d’organismes qui ne semblent appartenir à aucune des catégories de base. Les organismes que nous avons étudiés sont procaryotes en ce sens qu’ils n’ont pas de noyau et qu’ils ressemblent beaucoup à des bactéries ordinaires. Cependant, dans leur biochimie et dans la structure de certaines grosses molécules, ils sont aussi différents des autres procaryotes que des eucaryotes. Phylogénétiquement, ils ne sont ni procaryotes ni eucaryotes. Ils constituent un nouveau « royaume primaire », avec un statut complètement différent dans l’histoire et l’ordre naturel de la vie.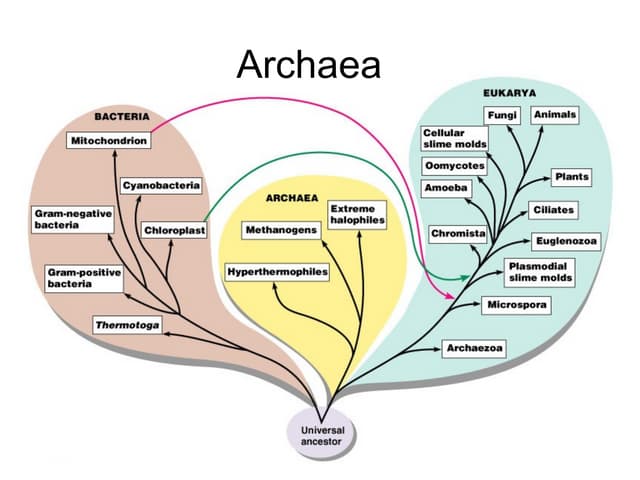 Nous avons nommé ces organismes archaebactéries. Le nom reflète une conjecture non testée sur leur statut évolutif. Les preuves phylogénétiques suggèrent que les archaebactéries sont au moins aussi anciennes que les autres grands groupes. De plus, certaines des archaebactéries ont une forme de métabolisme qui semble particulièrement bien adaptée aux conditions supposées avoir régné dans l’histoire primitive de la vie sur terre. Il semble donc possible que le groupe d’organismes le plus récent soit en fait le plus ancien.
Nous avons nommé ces organismes archaebactéries. Le nom reflète une conjecture non testée sur leur statut évolutif. Les preuves phylogénétiques suggèrent que les archaebactéries sont au moins aussi anciennes que les autres grands groupes. De plus, certaines des archaebactéries ont une forme de métabolisme qui semble particulièrement bien adaptée aux conditions supposées avoir régné dans l’histoire primitive de la vie sur terre. Il semble donc possible que le groupe d’organismes le plus récent soit en fait le plus ancien.
Le dossier évolutif

La Terre a quatre milliards et demi d’années et, d’après les archives fossiles macroscopiques, elle semble avoir été habitée pendant moins d’un septième de cette époque : toute la progression évolutive depuis les formes marines les plus anciennes jusqu’à l’homme ne s’étend que sur 600 millions d’années. Les empreintes fossiles d’organismes unicellulaires trop petits pour être vus à l’œil nu racontent une autre histoire. Les microfossiles de bactéries en particulier sont abondants dans les sédiments de tous âges ; ils ont été trouvés dans les plus anciennes roches sédimentaires intactes connues, des dépôts vieux de 3,5 milliards d’années en Australie. Au cours d’une énorme période de temps, pendant laquelle aucune forme supérieure n’existait, les bactéries sont apparues et ont rayonné pour former une grande variété de types habitant un grand nombre de niches écologiques. Jusqu’à récemment, cependant, on ne savait presque rien sur l’âge des micro-organismes. Les microfossiles bactériens sont des structures peu informatives ; peu de choses peuvent être déduites de l’empreinte d’une petite sphère ou d’une tige. Les principales indications paléontologiques de la nature des premières bactéries ne proviennent pas des microfossiles individuels mais des structures macroscopiques appelées stromatolites, que l’on pense être des tapis bactériens fossilisés : des colonies de bactéries incrustées de minéraux. Aujourd’hui, ces structures sont formées principalement par plusieurs types de bactéries photosynthétiques, généralement les cyanobactéries. Les stromatolithes fossilisés récemment ressemblent aux anciens à tel point qu’il semble tout à fait raisonnable de penser que les anciennes structures ont également été fabriquées par des bactéries photosynthétiques. Par conséquent, au moins certaines des anciennes bactéries doivent avoir été des photosynthétiseurs. En dehors de ce fait, pratiquement rien n’a pu être établi avec certitude sur les premiers micro-organismes. L’ensemble de l’arbre évolutif des bactéries est resté obscur, tout comme la base de l’arbre des formes de vie supérieures.
Jusqu’à récemment, cependant, on ne savait presque rien sur l’âge des micro-organismes. Les microfossiles bactériens sont des structures peu informatives ; peu de choses peuvent être déduites de l’empreinte d’une petite sphère ou d’une tige. Les principales indications paléontologiques de la nature des premières bactéries ne proviennent pas des microfossiles individuels mais des structures macroscopiques appelées stromatolites, que l’on pense être des tapis bactériens fossilisés : des colonies de bactéries incrustées de minéraux. Aujourd’hui, ces structures sont formées principalement par plusieurs types de bactéries photosynthétiques, généralement les cyanobactéries. Les stromatolithes fossilisés récemment ressemblent aux anciens à tel point qu’il semble tout à fait raisonnable de penser que les anciennes structures ont également été fabriquées par des bactéries photosynthétiques. Par conséquent, au moins certaines des anciennes bactéries doivent avoir été des photosynthétiseurs. En dehors de ce fait, pratiquement rien n’a pu être établi avec certitude sur les premiers micro-organismes. L’ensemble de l’arbre évolutif des bactéries est resté obscur, tout comme la base de l’arbre des formes de vie supérieures.
En reconstituant les premiers événements évolutifs, cependant, les biologistes ne se limitent pas aux archives fossiles géologiques. La cellule elle-même conserve des traces de son passé dans les séquences d’acides aminés de ses protéines et dans les séquences nucléotidiques de ses acides nucléiques : ADN et ARN. Ces archives vivantes sont potentiellement beaucoup plus riches et plus étendues que les archives fossiles, et elles remontent dans le temps au-delà des fossiles les plus anciens, jusqu’à la période où l’ancêtre commun de toute vie existait.
Afin de lire le dossier biochimique, il était nécessaire de développer une technologie pour déterminer (au moins en partie) la séquence d’un gène ou de l’ARN ou du produit protéique codé par un gène. Pour les protéines, cela est possible depuis environ 25 ans, mais le séquençage direct de l’ADN et de l’ARN n’est réalisable que depuis environ cinq ans. La nouvelle technologie de séquençage des acides nucléiques devrait permettre aux biologistes de découvrir dans un délai relativement court beaucoup plus sur l’histoire de la vie qu’on ne l’aurait cru possible. C’est en appliquant les techniques de séquençage au problème séculaire des relations naturelles entre bactéries que mes collègues et moi avons reconnu les archaebactéries comme une troisième forme de vie.
Eucaryotes et procaryotes
Afin d’apprécier le statut particulier des archaebactéries, il est utile de considérer certaines des caractéristiques déterminantes des eucaryotes et des procaryotes. La cellule eucaryote est relativement grande : environ 10 micromètres de côté. Il est entouré d’une double membrane, à l’intérieur de laquelle on distingue un certain nombre de structures elles-mêmes définies par des membranes. Le noyau contient la majeure partie du matériel génétique de la cellule. Les mitochondries en forme de bâtonnets sont le site de la respiration cellulaire, qui génère la principale monnaie énergétique de la cellule, l’adénosine triphosphate (ATP). Dans les cellules végétales, le chloroplaste, un autre corps en forme de bâtonnet, convertit l’énergie lumineuse en énergie chimique de l’ATP.
D’autres structures spécialisées telles que l’appareil de Golgi (pour la sécrétion) et les cils (pour la motilité) sont souvent présentes. De nombreuses cellules eucaryotes sont dotées d’un système membranaire, le réticulum endoplasmique, qui fournit une surface sur laquelle des réactions importantes telles que la synthèse de protéines ont lieu. La cellule procaryote est très différente. Il est généralement beaucoup plus petit que la cellule eucaryote : d’un facteur 10 en mesure linéaire et donc d’un facteur 1 000 en volume. La cellule procaryote est elle aussi circonscrite par une double membrane, et de plus elle a presque toujours une paroi cellulaire rigide. En revanche, aucune des structures internes caractéristiques de la cellule eucaryote n’est présente ; il n’y a pas de mitochondries ou de chloroplastes et bien sûr il n’y a pas de noyau lié à la membrane. Le génome – le complément total du matériel génétique – est limité à entre 2 000 et 3 000 gènes dans une cellule procaryote ; le génome eucaryote typique est plus grand de plusieurs ordres de grandeur.
La distinction entre eucaryotes et procaryotes a été initialement définie en termes de structures subcellulaires visibles au microscope. À ce niveau, toutes les cellules semblaient être soit grandes et complexes, et donc eucaryotes, soit petites et simples, et donc procaryotes. La distinction entre les deux types de cellules a finalement été portée au niveau biologique le plus fondamental, le niveau des molécules. Ici, les cellules eucaryotes et procaryotes ont de nombreuses caractéristiques en commun. Par exemple, ils traduisent l’information génétique en protéines selon le même code génétique. Même lorsque les processus moléculaires sont les mêmes, cependant, les détails des deux formes sont différents ; ils sont soit typiquement eucaryotes, soit typiquement procaryotes. Par exemple, les séquences d’acides aminés de diverses enzymes tendent à être typiquement procaryotes ou eucaryotes.
Cette conclusion a été tirée trop hâtivement ; l’attrait esthétique d’une dichotomie était trop grand. Simplement parce qu’il existe deux types de cellules au niveau microscopique, il ne s’ensuit pas qu’il doive y avoir seulement deux types au niveau moléculaire. La relation évolutive des procaryotes et des eucaryotes est en fait plus compliquée que les preuves citées ci-dessus ne l’indiquent. Deux organites eucaryotes, la mitochondrie et le chloroplaste, possèdent chacun leur propre ADN. De plus, les pigments du chloroplaste (les chlorophylles et les caroténoïdes) sont similaires à ceux des cyanobactéries. Les mitochondries et les chloroplastes ont la taille d’une bactérie leur appareil pour traduire l’information génétique en protéines diffère de l’appareil propre de la cellule eucaryote et a un certain nombre de propriétés en commun avec celui des procaryotes.
Ces faits et d’autres ont conduit à l’idée que les mitochondries et les chloroplastes descendent de procaryotes qui ont été piégés dans une cellule plus grande et ont établi une relation endosymbiotique avec elle. On pense que la mitochondrie était une bactérie respirante et que le chloroplaste était un parent photosynthétique des cyanobactéries. Cette conjecture, qui dans sa forme la plus simple a plus d’un siècle, a été essentiellement prouvée dans le cas du chloroplaste par la démonstration que la séquence nucléotidique de l’un des types d’ARN de l’organelle, l’ARN ribosomal, est spécifiquement liée à l’ARN ribosomal. -Séquences d’ARN chez les cyanobactéries. De même, l’ARN ribosomique de la mitochondrie chez les plantes semble être de type bactérien. Ainsi, il semble qu’au moins deux lignées de descendance procaryote soient représentées dans la cellule eucaryote.
L’urcaryote
Logiquement, la question suivante est : d’où vient le reste de la cellule eucaryote ? Quelle était la cellule hôte d’origine : l’urcaryote ? Il est généralement admis que la majeure partie de la cellule eucaryote (le noyau et les structures cytoplasmiques) représente une lignée distincte. La nature exacte de la cellule ancestrale n’est pas claire.  La plupart des chercheurs ont eu tendance à croire que la principale lignée eucaryote provenait également des bactéries ordinaires. L’idée est que certaines bactéries anaérobies tirant leur énergie de la fermentation des nutriments (plutôt que de leur oxydation) ont à un moment donné perdu leur paroi cellulaire résistante. Des organismes de ce genre sont connus ; ce sont les mycoplasmes. Une souche de mycoplasme a ensuite développé la capacité d’engloutir d’autres organismes, une capacité conservée par de nombreux eucaryotes aujourd’hui. Parmi les nombreux types d’organismes qu’un tel mycoplasme aurait pu ingérer, deux semblent avoir établi une relation endosymbiotique stable avec leur hôte et être devenus la mitochondrie et le chloroplaste. C’est ainsi que la cellule eucaryote est née. (L’origine de sa caractéristique déterminante, le noyau lié à la membrane, est encore loin d’être claire.)
La plupart des chercheurs ont eu tendance à croire que la principale lignée eucaryote provenait également des bactéries ordinaires. L’idée est que certaines bactéries anaérobies tirant leur énergie de la fermentation des nutriments (plutôt que de leur oxydation) ont à un moment donné perdu leur paroi cellulaire résistante. Des organismes de ce genre sont connus ; ce sont les mycoplasmes. Une souche de mycoplasme a ensuite développé la capacité d’engloutir d’autres organismes, une capacité conservée par de nombreux eucaryotes aujourd’hui. Parmi les nombreux types d’organismes qu’un tel mycoplasme aurait pu ingérer, deux semblent avoir établi une relation endosymbiotique stable avec leur hôte et être devenus la mitochondrie et le chloroplaste. C’est ainsi que la cellule eucaryote est née. (L’origine de sa caractéristique déterminante, le noyau lié à la membrane, est encore loin d’être claire.)
Ce point de vue est satisfaisant à certains égards, mais il n’explique pas les nombreuses différences entre les eucaryotes et les procaryotes. En particulier, il ne tient pas compte des différents détails des processus moléculaires ou des grandes différences dans les séquences d’acides aminés de protéines fonctionnellement analogues dans les deux types de cellules. Il est souvent pris pour acquis que les différences ne sont que la conséquence des nombreux petits changements dans la conception cellulaire qui seraient nécessaires pour passer de la condition procaryote simple à la condition eucaryote plus complexe. Il est douteux que tant de changements (changements dans la composition de presque toutes les enzymes, par exemple) puissent raisonnablement être expliqués de cette manière.
Essentiellement pour cette raison, certains biologistes pensent que la lignée qui a donné naissance à l’espèce urcaryote putative peut avoir divergé de la lignée procaryote à un moment antérieur, avant que l’ancêtre de la bactérie ne soit lui-même apparu. L’urcaryote aurait alors pu évoluer indépendamment vers une forme comparable en complexité à celle de la bactérie. Une telle hypothèse donnerait au moins plus de temps pour que des différences apparaissent entre les procaryotes et l’urcaryote. L’urcaryote représenterait alors une descendance distincte de celle des procaryotes, conformément à la dichotomie phylogénétique de base.
Il en était ainsi au début des années 1970. La phylogénie des eucaryotes supérieurs, couvrant quelque 500 millions d’années, était raisonnablement bien comprise, à l’exception de la jonction très importante des principales branches eucaryotes. Il y avait une hypothèse précise et largement acceptée concernant la manière dont la cellule eucaryote avait évolué. Les tests de l’hypothèse et les réponses aux questions restantes, cependant, résident dans les recoins inexplorés de la phylogénie bactérienne, à l’âge des micro-organismes.
Séquençage génétique

Les bactéries constituent un monde d’une variété extraordinaire, bien plus que ce que le microscope révèle. Les niches écologiques dans lesquelles ils se trouvent dépassent de loin en variété celles occupées par les formes de vie supérieures. Pendant un siècle, les microbiologistes ont tenté en vain de comprendre les relations naturelles entre les bactéries et d’imposer un ordre à l’étonnant éventail de formes, de physiologies et d’écologies. La variété parmi les bactéries est principalement la variété dans la simplicité, et elle fournit donc peu d’informations sur les relations phylogénétiques. Dans les organismes supérieurs, l’œil, par exemple, a évolué un certain nombre de fois, mais l’œil est suffisamment compliqué pour que les exemples évolués indépendamment se distinguent facilement les uns des autres.  Tel n’est généralement pas le cas pour la forme et la structure des bactéries ; bâtonnets, sphères et spirales, qui sont les formes bactériennes typiques, sont faciles à atteindre et ont évolué de nombreuses fois. Le même principe s’applique à la biochimie bactérienne. Bien que certaines caractéristiques bactériennes soient des indicateurs phylogénétiques valables, il est impossible de dire à l’avance lesquelles le sont et lesquelles ne le sont pas.
Tel n’est généralement pas le cas pour la forme et la structure des bactéries ; bâtonnets, sphères et spirales, qui sont les formes bactériennes typiques, sont faciles à atteindre et ont évolué de nombreuses fois. Le même principe s’applique à la biochimie bactérienne. Bien que certaines caractéristiques bactériennes soient des indicateurs phylogénétiques valables, il est impossible de dire à l’avance lesquelles le sont et lesquelles ne le sont pas.
La façon la plus simple dont la cellule est un enregistrement de son passé est en termes de séquences génétiques. Chaque gène qui existe dans une cellule aujourd’hui est une copie d’un gène qui existait il y a des éons. Ce n’est pas une copie exacte car des mutations ont altéré la séquence génétique d’origine, mais des vestiges de l’état d’origine persistent souvent. Ce qui fait d’un gène un superbe enregistrement du passé, c’est sa simplicité (il s’agit d’un réseau linéaire) et le fait que « l’espace » des séquences génétiques est énorme, de sorte que sur toute la durée de l’évolution, seule une petite fraction des séquences génétiques possibles peut ne jamais être réalisée. Ainsi, si deux gènes sont similaires sur une étendue comprenant un nombre significatif de nucléotides, cela ne peut signifier qu’ils ont un ancêtre en commun ; de telles molécules génétiquement apparentées sont dites homologues.
Une séquence génétique donne trois types d’informations évolutives. La séquence peut révéler des relations généalogiques, elle peut mesurer le temps d’évolution et c’est un enregistrement de caractéristiques ancestrales. Dans la mesure où deux gènes pour la même fonction dans différents organismes sont liés, les organismes sont liés. La mesure dans laquelle deux de ces séquences diffèrent mesure le temps écoulé depuis que les organismes ont divergé d’un ancêtre commun. À partir d’un vaste ensemble de séquences apparentées, on peut construire un arbre phylogénétique dans lequel les points de ramification mesurent (approximativement) les temps relatifs des bifurcations. Enfin, des comparaisons de fils parmi un vaste ensemble de séquences homologues permettent de reconstituer avec une certaine précision diverses versions ancestrales d’un gène.
Étant donné que la relation entre un gène et son produit (soit une protéine, soit l’un des nombreux types de molécules d’ARN) est généralement colinéaire, la séquence du produit est généralement aussi utile pour les études phylogénétiques que la séquence du gène lui-même. Parce que jusqu’à récemment seules les protéines pouvaient être séquencées, c’est par des comparaisons de protéines que les premières phylogénies basées sur l’évolution moléculaire ont été construites. Les comparaisons de la protéine respiratoire cytochrome c s’avèrent particulièrement précieuses pour confirmer et étendre l’arbre phylogénétique des organismes supérieurs. 
Méthanogènes
En 1977, l’identification des méthanogènes, une forme de vie datant d’environ 3,5 milliards d’années, a été rapportée par des scientifiques de l’Université et de l’Illinois. Le microbiologiste Carl R. Woese -1928-2012) a longtemps étudié la voie évolutive de l’ADN et de l’ARN. En 1976, il est approché par son collègue Ralph Wolfe, qui présente un groupe d’organismes producteurs de méthane. Woese a étudié leur ARN et a reconnu leur manque de séquences oligonucléotidiques entières. Il a découvert que les organismes étaient si différents des bactéries qu’ils méritaient leur propre branche de l’arbre généalogique en tant que troisième domaine de la vie, Archaea. Les méthanogènes se trouvent dans des environnements pauvres en oxygène et obtiennent principalement leur énergie en réduisant le dioxyde de carbone et en oxydant l’hydrogène, avec la production de méthane.
https://www.youtube.com/watch?v=9pvSZTQTlc0
https://annalsmicrobiology.biomedcentral.com/articles/10.1007/s13213-009-0008-5
https://link.springer.com/referenceworkentry/10.1007/0-387-30743-5_9
https://www.scientificamerican.com/article/archaebacteria-the-third/
https://link.springer.com/chapter/10.1007/978-3-662-13187-9_76