 Robert F. Furchgott, pharmacologue et éducateur américain (co-lauréat du prix Nobel de physiologie ou médecine, 1998)
Robert F. Furchgott, pharmacologue et éducateur américain (co-lauréat du prix Nobel de physiologie ou médecine, 1998) Lauréat du prix Nobel, Robert F. Furchgott (1916–2009)
Lauréat du prix Nobel, Robert F. Furchgott (1916–2009) Robert F. Furchgott, pharmacologue et co-lauréat du prix Nobel de médecine ou de physiologie (1998) est décédé le 12 mai 2009 à l’âge de 92 ans. En débloquant les actions biologiques étonnamment diverses de l’oxyde nitrique, Furchgott laisse derrière lui un riche héritage qui a à la fois révolutionné notre compréhension de la physiologie humaine et stimulé de nouvelles opportunités passionnantes pour le développement de médicaments dans un large éventail de conditions pathologiques. Dans cet article, William Martin, qui a travaillé avec Furchgott pendant 2 ans (1983-1985), suite à la découverte passionnante du facteur relaxant/oxyde nitrique dérivé de l’endothélium, rend hommage à son ami proche et collègue.
Robert F. Furchgott, pharmacologue et co-lauréat du prix Nobel de médecine ou de physiologie (1998) est décédé le 12 mai 2009 à l’âge de 92 ans. En débloquant les actions biologiques étonnamment diverses de l’oxyde nitrique, Furchgott laisse derrière lui un riche héritage qui a à la fois révolutionné notre compréhension de la physiologie humaine et stimulé de nouvelles opportunités passionnantes pour le développement de médicaments dans un large éventail de conditions pathologiques. Dans cet article, William Martin, qui a travaillé avec Furchgott pendant 2 ans (1983-1985), suite à la découverte passionnante du facteur relaxant/oxyde nitrique dérivé de l’endothélium, rend hommage à son ami proche et collègue.
 Robert F. Furchgottétait un biochimiste américain qui a découvert que l’oxyde nitrique (NO) agit comme une molécule de signalisation dans le système cardiovasculaire des mammifères. Cette découverte lui a valu une part du prix Nobel de physiologie ou médecine en 1998 ainsi que plusieurs autres prix et distinctions prestigieux. Fils d’un propriétaire de grand magasin, il a passé de nombreuses années de sa jeunesse dans de petites villes où il a développé un amour pour la nature et les sciences. Enfant, il aimait visiter les plages, les marais et les bois, et était un passionné d’ornithologie. Dès son plus jeune âge, il n’y avait aucun doute dans son esprit qu’il était destiné à devenir un scientifique et les encouragements de ses parents ont encore alimenté ses ambitions. Après avoir obtenu un diplôme en chimie de l’Université de Caroline du Nord à Chapel Hill, il a obtenu son doctorat en biochimie de l’Université Northwestern.
Robert F. Furchgottétait un biochimiste américain qui a découvert que l’oxyde nitrique (NO) agit comme une molécule de signalisation dans le système cardiovasculaire des mammifères. Cette découverte lui a valu une part du prix Nobel de physiologie ou médecine en 1998 ainsi que plusieurs autres prix et distinctions prestigieux. Fils d’un propriétaire de grand magasin, il a passé de nombreuses années de sa jeunesse dans de petites villes où il a développé un amour pour la nature et les sciences. Enfant, il aimait visiter les plages, les marais et les bois, et était un passionné d’ornithologie. Dès son plus jeune âge, il n’y avait aucun doute dans son esprit qu’il était destiné à devenir un scientifique et les encouragements de ses parents ont encore alimenté ses ambitions. Après avoir obtenu un diplôme en chimie de l’Université de Caroline du Nord à Chapel Hill, il a obtenu son doctorat en biochimie de l’Université Northwestern. 
Grands travaux : Il est connu pour ses recherches sur l’oxyde nitrique (NO) qui ont conduit à la découverte que le NO agit comme une molécule de signalisation dans le système cardiovasculaire des mammifères, ce qui est l’une des découvertes les plus importantes de l’histoire de la médecine cardiovasculaire. Son travail a également ouvert la voie à d’autres recherches qui ont conduit au développement du Viagra, le médicament anti-impuissance.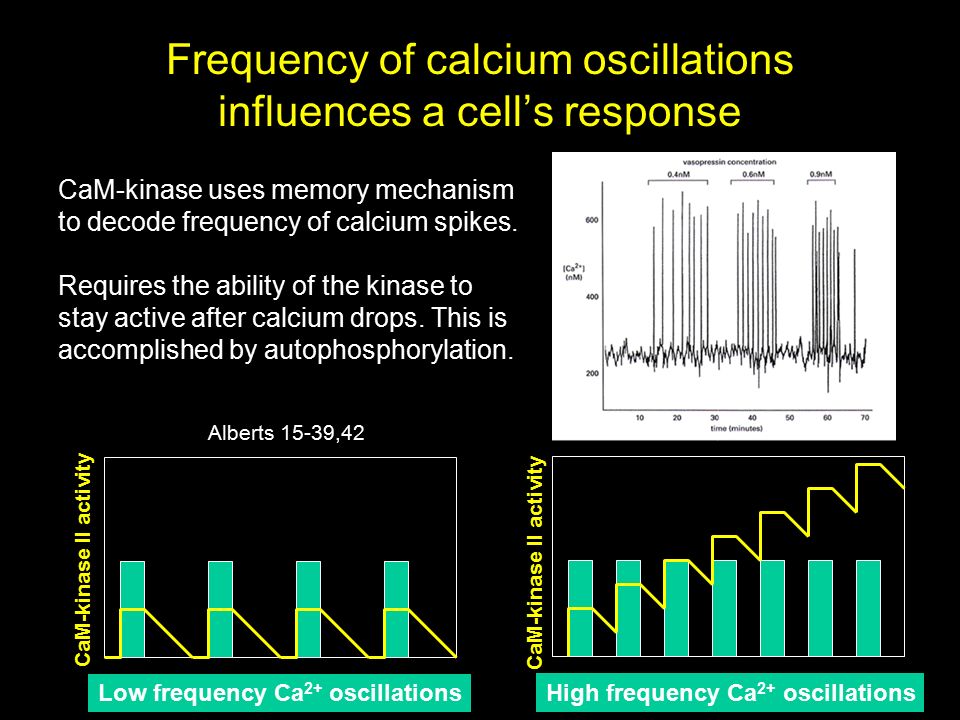 Récompenses et réalisations : Furchgott a reçu plusieurs prix prestigieux, dont le prix Roussel-Uclaf pour la recherche en transduction du signal (1993) et la médaille d’or Wellcome, British Pharmacological Society (1995). En 1996, Furchgott et Ferid Murad ont reçu le prix Albert Lasker pour la recherche médicale fondamentale.Robert F. Furchgott avec Louis J. Ignarro et Ferid Murad ont reçu conjointement le prix Nobel de physiologie ou médecine 1998 « pour leurs découvertes concernant l’oxyde nitrique en tant que molécule de signalisation dans le système cardiovasculaire ».
Récompenses et réalisations : Furchgott a reçu plusieurs prix prestigieux, dont le prix Roussel-Uclaf pour la recherche en transduction du signal (1993) et la médaille d’or Wellcome, British Pharmacological Society (1995). En 1996, Furchgott et Ferid Murad ont reçu le prix Albert Lasker pour la recherche médicale fondamentale.Robert F. Furchgott avec Louis J. Ignarro et Ferid Murad ont reçu conjointement le prix Nobel de physiologie ou médecine 1998 « pour leurs découvertes concernant l’oxyde nitrique en tant que molécule de signalisation dans le système cardiovasculaire ».
Biographique Robert F. Furchgott (1916-2009); Le prix Nobel de physiologie ou médecine 1998
 Au cours des deux premières années du lycée, j’ai su que j’aimerais être scientifique. Mes parents étaient encourageants : ils m’ont offert des trousses de chimie et un petit microscope comme cadeaux. J’aimais lire des livres populaires sur les scientifiques, même s’il n’y en avait pas beaucoup à l’époque. Mon père était abonné au Sunday New York Times, dans lequel il y avait souvent une chronique sur la science que je trouvais très excitante.
Au cours des deux premières années du lycée, j’ai su que j’aimerais être scientifique. Mes parents étaient encourageants : ils m’ont offert des trousses de chimie et un petit microscope comme cadeaux. J’aimais lire des livres populaires sur les scientifiques, même s’il n’y en avait pas beaucoup à l’époque. Mon père était abonné au Sunday New York Times, dans lequel il y avait souvent une chronique sur la science que je trouvais très excitante.
Pendant les quatre années où j’étais au lycée, mon frère aîné Arthur était à l’Université de Caroline du Nord à Chapel Hill. Je voulais aussi aller à l’université là-bas, mais cela n’a pas été possible lorsque j’ai terminé le lycée en 1933 car les frais de scolarité pour moi, en tant que résident hors de l’État, étaient plus que ce que mon père pouvait se permettre à l’époque. J’ai donc passé ma première année à l’Université de Caroline du Sud, où mes frais de scolarité étaient beaucoup moins élevés. Cependant, à l’été 1934, mon père a déménagé son entreprise d’Orangeburg à Goldsboro, en Caroline du Nord, où il a estimé que l’économie locale était meilleure. Alors maintenant, en tant que résident de Caroline du Nord, j’ai pu m’inscrire à l’Université de Chapel Hill en deuxième année avec une spécialisation en chimie.

Northwestern et Cold Spring Harbor (1937-1940)
Avant d’aller à Chicago, j’ai travaillé pendant deux mois d’été en 1937 pour Eastern Airlines à l’aéroport de Philadelphie – un emploi que mon frère aîné Arthur, qui était employé par cette compagnie aérienne, m’a aidé à obtenir. Le travail m’a permis d’économiser de l’argent et m’a également permis de voyager gratuitement en avion jusqu’à Chicago. Cela m’a beaucoup aidé puisque mon salaire en tant qu’assistant d’enseignement à Northwestern n’était que de 50 $ par mois pour une année universitaire de neuf mois. Quand je suis arrivé à Chicago, il avait déjà été prévu que je partage une chambre avec deux étudiants diplômés plus avancés. Vivre à Chicago était bien différent de vivre dans les Carolines. Quand je me rendais au travail à pied l’hiver depuis notre maison de chambres, qui se trouvait à environ un mile de l’école de médecine, le vent froid soufflant du lac Michigan le long de Chicago Avenue était toute une expérience pour un garçon du Sud.
Mon travail de cours à Northwestern était en partie à la faculté de médecine et en partie sur le campus d’Evanston où je voyageais via l’El. Au campus d’Evanston, mes cours étaient principalement en chimie physique sous la direction du Dr Malcolm Dole, qui faisait également partie de mon comité consultatif de doctorat. Sur le campus de Chicago, j’ai dû suivre la physiologie et la bactériologie (avec des étudiants en médecine), le cours de Henry Bull sur la chimie physique en biochimie et quelques cours de deuxième cycle en physiologie et biochimie. Le cours de physiologie était sous la direction du Dr Andrew Ivy, qui avait constitué une importante faculté de département de physiologie pour l’époque. En revanche, la faculté de biochimie ne comprenait que le président, le Dr Chester Farmer, le Dr Bull et deux chargés de cours à temps partiel. Mon travail en laboratoire avec Bull a commencé par la préparation d’albumine d’œuf purifiée. Il étudiait les changements physico-chimiques de cette protéine après différentes méthodes de dénaturation. Il avait commencé à m’impliquer dans certaines de ses études lorsque l’été 1938 arriva, et cela s’avéra être un été spécial pour moi. Bull avait été invité à présenter un article sur ses travaux au sixième Symposium de Cold Spring Harbor sur la biologie quantitative qui devait avoir lieu au Cold Spring Harbor Biological Laboratory de la Long Island Biological Association. Le thème du symposium, qui devait se dérouler tranquillement pendant cinq semaines, était la structure et la fonction des protéines. Bull avait obtenu la permission du directeur du Cold Spring Harbor Laboratory, le Dr Eric Ponder, pour que j’assiste au symposium, tout en gagnant ma chambre et ma pension en faisant fonctionner le projecteur de diapositives à lanterne lors des conférences. Le symposium était très excitant. J’ai rencontré de nombreux scientifiques distingués. Ponder et un médecin-chercheur, Harold Abramson, se sont arrangés pour que je participe à un projet de recherche au laboratoire pour le reste de l’été après la fin du symposium. Le projet portait sur la mobilité électrophorétique des érythrocytes et des fantômes de lapin, mesurée à l’aide d’une cellule de microélectrophorèse et d’une microscopie à fond clair et noir.
Mon travail en laboratoire avec Bull a commencé par la préparation d’albumine d’œuf purifiée. Il étudiait les changements physico-chimiques de cette protéine après différentes méthodes de dénaturation. Il avait commencé à m’impliquer dans certaines de ses études lorsque l’été 1938 arriva, et cela s’avéra être un été spécial pour moi. Bull avait été invité à présenter un article sur ses travaux au sixième Symposium de Cold Spring Harbor sur la biologie quantitative qui devait avoir lieu au Cold Spring Harbor Biological Laboratory de la Long Island Biological Association. Le thème du symposium, qui devait se dérouler tranquillement pendant cinq semaines, était la structure et la fonction des protéines. Bull avait obtenu la permission du directeur du Cold Spring Harbor Laboratory, le Dr Eric Ponder, pour que j’assiste au symposium, tout en gagnant ma chambre et ma pension en faisant fonctionner le projecteur de diapositives à lanterne lors des conférences. Le symposium était très excitant. J’ai rencontré de nombreux scientifiques distingués. Ponder et un médecin-chercheur, Harold Abramson, se sont arrangés pour que je participe à un projet de recherche au laboratoire pour le reste de l’été après la fin du symposium. Le projet portait sur la mobilité électrophorétique des érythrocytes et des fantômes de lapin, mesurée à l’aide d’une cellule de microélectrophorèse et d’une microscopie à fond clair et noir.
À la fin de l’été, je m’intéressais beaucoup à la chimie physique de la membrane des globules rouges. Quand je suis retourné à Northwestern à l’automne 1938, Bull a approuvé la poursuite de mes recherches sur les globules rouges en tant que projet de thèse de doctorat. En particulier, j’étais fasciné par le phénomène inexpliqué de la transformation des globules rouges de mammifères, suspendus dans une solution saline isotonique non tamponnée à partir de disques en sphères parfaites lorsqu’une petite goutte de la suspension était placée entre la lame et la lamelle. J’ai découvert que la transformation disque-sphère dépendait de deux facteurs. Le premier était une augmentation du pH à plus de 9,0 dans la suspension non tamponnée, en raison de la nature alcaline des surfaces de verre (le pH étant mesuré avec une électrode en verre semi-micro que j’ai construite). Le deuxième facteur était l’élimination de la suspension des globules rouges par adsorption sur les surfaces en verre de la lame et de la lamelle couvre-objet d’une substance dans la suspension qui empêchait la formation de sphères lors de l’élévation du pH de la suspension. J’ai démontré que cette substance, que j’ai appelée le facteur anti-sphère, était de l’albumine sérique qui ne pouvait pas être efficacement éliminée des globules rouges simplement par de multiples lavages et centrifugations. En plus des travaux sur les changements de forme des érythrocytes, mon travail de thèse de doctorat a impliqué des études supplémentaires sur l’électrophorèse des cellules dans diverses conditions et sur d’autres aspects de la chimie physique des membranes érythrocytaires.  J’ai démontré que cette substance, que j’ai appelée le facteur anti-sphère, était de l’albumine sérique qui ne pouvait pas être efficacement éliminée des globules rouges simplement par de multiples lavages et centrifugations. En plus des travaux sur les changements de forme des érythrocytes, mon travail de thèse de doctorat a impliqué des études supplémentaires sur l’électrophorèse des cellules dans diverses conditions et sur d’autres aspects de la chimie physique des membranes érythrocytaires. J’ai démontré que cette substance, que j’ai appelée le facteur anti-sphère, était de l’albumine sérique qui ne pouvait pas être efficacement éliminée des globules rouges simplement par de multiples lavages et centrifugations. En plus des travaux sur les changements de forme des érythrocytes, mon travail de thèse de doctorat a impliqué des études supplémentaires sur l’électrophorèse des cellules dans diverses conditions et sur d’autres aspects de la chimie physique des membranes érythrocytaires.
J’ai démontré que cette substance, que j’ai appelée le facteur anti-sphère, était de l’albumine sérique qui ne pouvait pas être efficacement éliminée des globules rouges simplement par de multiples lavages et centrifugations. En plus des travaux sur les changements de forme des érythrocytes, mon travail de thèse de doctorat a impliqué des études supplémentaires sur l’électrophorèse des cellules dans diverses conditions et sur d’autres aspects de la chimie physique des membranes érythrocytaires. J’ai démontré que cette substance, que j’ai appelée le facteur anti-sphère, était de l’albumine sérique qui ne pouvait pas être efficacement éliminée des globules rouges simplement par de multiples lavages et centrifugations. En plus des travaux sur les changements de forme des érythrocytes, mon travail de thèse de doctorat a impliqué des études supplémentaires sur l’électrophorèse des cellules dans diverses conditions et sur d’autres aspects de la chimie physique des membranes érythrocytaires.
Au cours de l’été 1939, à l’invitation de Ponder, avec qui j’ai eu une longue correspondance au cours de l’année et qui était devenu en fait le principal conseiller pour mes recherches de thèse de doctorat, je suis retourné à Cold Spring Harbor pour poursuivre des recherches sur les globules rouges. Pour gagner ma chambre et ma pension, j’ai servi sur des tables dans la salle à manger commune. J’ai aussi pu assister aux conférences du symposium de cette année-là, qui portaient sur les oxydations biologiques. Là, j’ai pris conscience pour la première fois des nouveaux développements du métabolisme énergétique oxydatif et de l’importance des composés phosphatés à haute énergie. Parmi les nombreux biochimistes exceptionnels présents figuraient L. Michaelis, Fritz Lipmann et Carl Cori. Ponder et sa jeune épouse Ruth m’ont été très accueillants. J’ai été très impressionné par son habileté à appliquer les mathématiques à ses recherches, sa facilité en rédaction scientifique et sa vaste collection de disques de musique classique. J’ai pu terminer et défendre ma thèse à temps pour recevoir le doctorat. diplôme en juin 1940. Plus tôt ce printemps-là, j’avais assisté à la réunion annuelle de la Fédération des sociétés américaines de biologie expérimentale (FASEB) à la Nouvelle-Orléans. Heureusement, Henry Tauber, un biochimiste autrichien travaillant pour une société pharmaceutique à Chicago, m’avait demandé de partager la conduite de sa voiture lors de l’aller-retour vers la Nouvelle-Orléans ainsi que sa chambre dans un hôtel délabré de la Nouvelle-Orléans. Ainsi, j’ai pu assister à cette réunion à très peu de frais. Lors de la réunion du FASEB à la Nouvelle-Orléans, où les rassemblements de participants étaient encore appelés « fumeurs » et où même un repas raffiné ne coûtait pas plus de deux dollars, j’ai eu des entretiens avec des personnes sur d’éventuels emplois post-doctoraux. L’une des entrevues était avec le Dr Ephraim Shorr, professeur agrégé de médecine à la Cornell University Medical School de New York, que j’avais rencontré à Cold Spring Harbor l’été précédent. Quelques semaines plus tard, Shorr m’a proposé un poste postdoctoral dans son laboratoire. Bien que j’espérais obtenir un poste qui me permettrait de continuer à travailler sur la chimie physique des protéines ou des membranes cellulaires, aucun ne s’est présenté et j’ai accepté le poste chez Shorr, étant entendu que je commencerais en septembre.
J’ai pu terminer et défendre ma thèse à temps pour recevoir le doctorat. diplôme en juin 1940. Plus tôt ce printemps-là, j’avais assisté à la réunion annuelle de la Fédération des sociétés américaines de biologie expérimentale (FASEB) à la Nouvelle-Orléans. Heureusement, Henry Tauber, un biochimiste autrichien travaillant pour une société pharmaceutique à Chicago, m’avait demandé de partager la conduite de sa voiture lors de l’aller-retour vers la Nouvelle-Orléans ainsi que sa chambre dans un hôtel délabré de la Nouvelle-Orléans. Ainsi, j’ai pu assister à cette réunion à très peu de frais. Lors de la réunion du FASEB à la Nouvelle-Orléans, où les rassemblements de participants étaient encore appelés « fumeurs » et où même un repas raffiné ne coûtait pas plus de deux dollars, j’ai eu des entretiens avec des personnes sur d’éventuels emplois post-doctoraux. L’une des entrevues était avec le Dr Ephraim Shorr, professeur agrégé de médecine à la Cornell University Medical School de New York, que j’avais rencontré à Cold Spring Harbor l’été précédent. Quelques semaines plus tard, Shorr m’a proposé un poste postdoctoral dans son laboratoire. Bien que j’espérais obtenir un poste qui me permettrait de continuer à travailler sur la chimie physique des protéines ou des membranes cellulaires, aucun ne s’est présenté et j’ai accepté le poste chez Shorr, étant entendu que je commencerais en septembre.
La raison pour laquelle j’ai attendu jusqu’en septembre pour commencer à travailler à Cornell était que je voulais passer un été de plus au laboratoire de biologie de Cold Spring Harbor. Cette fois, cependant, j’y suis allé en tant que conférencier invité au symposium qui, cet été-là, portait sur le thème de la perméabilité des membranes cellulaires. Ma conférence s’intitulait « Observations sur la structure des fantômes de globules rouges ». Lors de ce symposium, il y avait encore un certain nombre de scientifiques distingués établis comme KS Cole, Robert Chambers et FO Schmitt ; et en plus, un certain nombre de jeunes scientifiques brillants comme Hans Neurath, qui avait également participé au symposium de 1938, Hugh Davson, qui avec Danielli avait développé le modèle de membrane bicouche lipidique, et Benjamin Zweifach, avec qui je devais collaborer plus tard dans la recherche .
Collège de médecine de l’Université Cornell (1940-1949)
Je suis resté au Cornell University Medical College travaillant dans le laboratoire d’Ephraim Shorr pendant neuf ans. Quand je suis arrivé, Sam Barker, un jeune associé de recherche, était là pour m’enseigner les méthodes et les procédures qu’ils utilisaient pour étudier le métabolisme des tissus (en grande partie à l’aide de manomètres Warburg) et le renouvellement de fractions de phosphate organique tissulaire plutôt mal définies du muscle cardiaque canin. lors des incubations in vitro.Pour de telles études, le laboratoire a été l’un des premiers à utiliser du phosphate radioactif, que nous avons obtenu du laboratoire du cyclotron de Berkeley. Barker est parti vers la fin de ma première année à Cornell ; et j’étais alors chargé de diriger le laboratoire de Shorr. Shorr lui-même participait parfois à la préparation de tissus pour les expériences de Warburg. Il était tout à fait capable en laboratoire en plus d’être un clinicien très occupé et excellent.
Au cours de mes deux premières années à Cornell, mon projet principal portait sur l’échange et le renouvellement du phosphate, en utilisant du phosphate radioactif et des tranches de muscle ventriculaire gauche de chien. Un article complet sur le travail a été publié dans la revue Biological Chemistry en 1943. Les méthodes et l’équipement que nous avons utilisés dans ce travail ont depuis longtemps été remplacés, mais nous avons réussi avec des méthodes chimiques et quelques premières méthodes enzymatiques pour montrer le chiffre d’affaires extrêmement rapide de la créatinine. phosphate et le phosphate terminal de l’ATP dans le muscle cardiaque au repos.
L’article de 1943 a été ma première publication complète après trois ans de travail à Cornell. L’une des raisons probables de la faible production était que les États-Unis étaient entrés dans la Seconde Guerre mondiale en décembre 1941 et que Shorr, comme beaucoup d’autres, a commencé à entreprendre des recherches plus pertinentes pour l’effort de guerre. Avec le soutien du gouvernement et d’autres sources, il a réorienté les principales recherches du laboratoire vers le choc circulatoire – d’abord sur les changements du métabolisme énergétique des tissus résultant de l’hypoxie associée au choc hémorragique, puis principalement sur les facteurs pouvant expliquer le choc «irréversible», la condition dans lequel rétablissement du volume sanguin n’est plus capable d’augmenter la pression et de maintenir la vie chez l’animal soumis à une pression sanguine basse maintenue à la suite d’une hémorragie contrôlée. Pour l’aider dans cette nouvelle ligne de recherche, Shorr a recruté Benjamin Zweifach, puis un jeune physiologiste brillant qui s’était formé avec Robert Chambers et avait mis au point une belle méthode d’observation microscopique du flux sanguin dans une partie du mésentère (la zone «mésoappendice») du rat anesthésié. En bref, le « test de méso-appendice de rat », mené par Zweifach et des techniciens qu’il a formés, a mis en évidence en 1944 deux facteurs vasoactifs dans le choc circulatoire. Le premier facteur est apparu dans le plasma des chiens au stade précoce réversible (par transfusion) de l’hémorragie.  Des injections intraveineuses de ce plasma ont augmenté la sensibilité des petites artérioles et des sphincters précapillaires à l’épinéphrine appliquée localement dans le test du mésoappendice. Ce facteur a été appelé VEM (pour matériaux vasoexcitateurs). Au fur et à mesure que le stade irréversible du choc circulatoire se développait, L’activité VEM a disparu du plasma et un nouveau facteur est apparu qui a nettement diminué la sensibilité à l’épinéphrine dans le test du mésoappendice. Ce facteur a été nommé VDM (pour matériel vasodépresseur). Nous avons développé des preuves, en partie à partir dedes expériences in vitro avec des tranches de tissu, que le rein hypoxique était la source probable de VEM et que le foie hypoxique était la source probable de VDM. À la fin de 1945, ces développements ont conduit à un article principal dans la revue Science par Shorr, Zweifach et moi-même.
Des injections intraveineuses de ce plasma ont augmenté la sensibilité des petites artérioles et des sphincters précapillaires à l’épinéphrine appliquée localement dans le test du mésoappendice. Ce facteur a été appelé VEM (pour matériaux vasoexcitateurs). Au fur et à mesure que le stade irréversible du choc circulatoire se développait, L’activité VEM a disparu du plasma et un nouveau facteur est apparu qui a nettement diminué la sensibilité à l’épinéphrine dans le test du mésoappendice. Ce facteur a été nommé VDM (pour matériel vasodépresseur). Nous avons développé des preuves, en partie à partir dedes expériences in vitro avec des tranches de tissu, que le rein hypoxique était la source probable de VEM et que le foie hypoxique était la source probable de VDM. À la fin de 1945, ces développements ont conduit à un article principal dans la revue Science par Shorr, Zweifach et moi-même.
Pendant les années de guerre, je n’étais pas uniquement impliqué dans la recherche sur le métabolisme tissulaire et le choc circulatoire. En 1943, Eugene DuBois, président du département de physiologie de Cornell, s’est arrangé pour que je rejoigne son département en tant qu’instructeur afin de remplacer un membre du personnel perdu au service militaire. Bien que j’enseignais en physiologie, je passais la plupart de mon temps à faire de la recherche dans le laboratoire de Shorr, qui était en partie financé par l’Office fédéral de la recherche scientifique et du développement. Les travaux sur VEM et VDM se sont poursuivis après la fin de la guerre. J’avais tenté d’isoler le matériau de type VEM qui s’accumulait dans le liquide d’incubation lorsque les tranches de rein étaient incubées en anaérobie. J’ai pu le concentrer quelque peu et il a semblé être un peptide dialysable labile, mais je n’ai pas réussi à l’isoler. D’autre part, Abraham Mazur, un professeur de biochimie au City College de New York qui travaillait à temps partiel avec nous, a purifié du foie un matériau de type VDM qui semblait être de la ferritine. (Ferritine ou non, nous pourrions maintenant nous demander si le VDM pourrait d’une manière ou d’une autre être lié à l’oxyde nitrique !)
Malheureusement, la seule procédure d’essai biologique pour détecter l’activité VEM et VDM était celle impliquant des changements de sensibilité à l’épinéphrine dans le test du mésoappendice chez le rat. Les injections intraveineuses de solutions contenant des taux élevés de VEM impur ou de ferritine purifiée n’ont pas eu d’effet sur la tension artérielle chez les animaux de laboratoire. Les tentatives de développement d’un système d’essai biologique in vitro ont également échoué. Ces échecs ont tempéré mon enthousiasme, et je pense celui de Zweifach, pour l’importance du VEM et du VDM dans la régulation de la circulation. Cependant, les tentatives infructueuses de développement d’un bioessai in vitro pour le VEM et le VDM ont été très importantes pour moi car elles m’ont initié à la pharmacologie du muscle lisse, un sujet qui m’intéresse depuis lors. Deux des préparations isolées de muscles lisses que j’ai testées sans succès pour le dosage biologique de VEM et VDM étaient une bande d’aorte de lapin coupée en hélice, qui répondait par une contraction à l’épinéphrine, et un segment longitudinal de duodénum de lapin, qui présentait des contractions rythmiques spontanées inhibées. par l’épinéphrine et stimulée par l’acétylcholine. À cette époque, les contractions de ces préparations de muscles lisses montées dans des bains d’organes étaient enregistrées avec des leviers isotoniques sur des kymographes. Un jour, au cours d’essais sur des segments de duodénum de lapin montés dans une solution de Krebs oxygénée, j’ai été surpris de voir que pendant les premières heures de l’expérience, l’amplitude de la contraction ne se stabilisait pas comme d’habitude mais diminuait progressivement et nettement même si la fréquence rythmique resté inchangé. Je soupçonnais que mon technicien avait oublié d’ajouter du glucose à la solution de Krebs. L’ajout de glucose a maintenant rapidement augmenté l’amplitude de la contraction jusqu’au niveau normal. Cette découverte a conduit à une procédure simple pour découvrir quels sucres et acides gras pourraient être utilisés comme énergie pour la contraction du muscle lisse intestinal dans des conditions aérobies et anaérobies et pour analyser les sites d’action des inhibiteurs métaboliques.
Deux des préparations isolées de muscles lisses que j’ai testées sans succès pour le dosage biologique de VEM et VDM étaient une bande d’aorte de lapin coupée en hélice, qui répondait par une contraction à l’épinéphrine, et un segment longitudinal de duodénum de lapin, qui présentait des contractions rythmiques spontanées inhibées. par l’épinéphrine et stimulée par l’acétylcholine. À cette époque, les contractions de ces préparations de muscles lisses montées dans des bains d’organes étaient enregistrées avec des leviers isotoniques sur des kymographes. Un jour, au cours d’essais sur des segments de duodénum de lapin montés dans une solution de Krebs oxygénée, j’ai été surpris de voir que pendant les premières heures de l’expérience, l’amplitude de la contraction ne se stabilisait pas comme d’habitude mais diminuait progressivement et nettement même si la fréquence rythmique resté inchangé. Je soupçonnais que mon technicien avait oublié d’ajouter du glucose à la solution de Krebs. L’ajout de glucose a maintenant rapidement augmenté l’amplitude de la contraction jusqu’au niveau normal. Cette découverte a conduit à une procédure simple pour découvrir quels sucres et acides gras pourraient être utilisés comme énergie pour la contraction du muscle lisse intestinal dans des conditions aérobies et anaérobies et pour analyser les sites d’action des inhibiteurs métaboliques.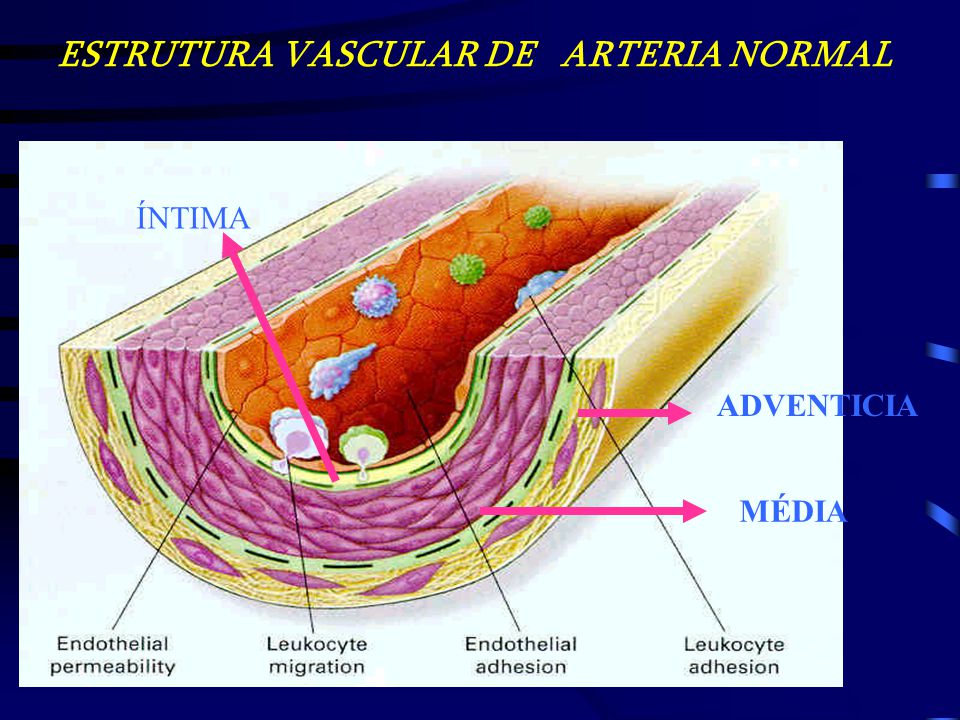 Au printemps 1949, j’ai eu deux offres intéressantes au niveau de professeur adjoint – une en physiologie à Duke et une en pharmacologie à la faculté de médecine de l’Université de Washington. J’ai choisi l’Université de Washington, en partie parce que le nouveau président, Oliver Lowry, était quelqu’un que j’avais connu au Enzyme Club de New York et en partie parce que j’avais commencé à m’intéresser beaucoup à la pharmacologie en tant que discipline. C’était en partie à cause des études que j’avais commencées sur les effets des médicaments et d’autres agents sur les préparations de muscles lisses in vitro, mais aussi en grande partie à cause de mon amitié étroite avec Walter Riker, qui était alors membre junior du département de pharmacologie de Cornell au début d’une brillante carrière. Son enthousiasme pour la recherche en pharmacologie était contagieux.
Au printemps 1949, j’ai eu deux offres intéressantes au niveau de professeur adjoint – une en physiologie à Duke et une en pharmacologie à la faculté de médecine de l’Université de Washington. J’ai choisi l’Université de Washington, en partie parce que le nouveau président, Oliver Lowry, était quelqu’un que j’avais connu au Enzyme Club de New York et en partie parce que j’avais commencé à m’intéresser beaucoup à la pharmacologie en tant que discipline. C’était en partie à cause des études que j’avais commencées sur les effets des médicaments et d’autres agents sur les préparations de muscles lisses in vitro, mais aussi en grande partie à cause de mon amitié étroite avec Walter Riker, qui était alors membre junior du département de pharmacologie de Cornell au début d’une brillante carrière. Son enthousiasme pour la recherche en pharmacologie était contagieux.
À l’été 1949, ma famille et moi avons conduit de New York à Saint-Louis. Ma femme, Lenore, originaire de New York, a dit qu’elle avait l’impression d’aller vers l’Ouest dans un chariot couvert. À cette époque, nous avions deux filles, âgées de quatre et un ans. Plus tard, nous avons eu une troisième fille née à Saint-Louis. On peut noter ici qu’aucune de mes filles n’est devenue scientifique. Au lieu de cela, ils sont tous allés dans l’art (comme mon jeune frère, Max). On peut également noter ici que ma femme Lenore est décédée en 1983 ; et que maintenant j’ai une nouvelle femme, Margaret (Maggie). J’ai eu beaucoup de chance d’avoir des épouses qui ont encouragé mon travail, même si cela a souvent réduit le temps que je pouvais consacrer aux affaires familiales. 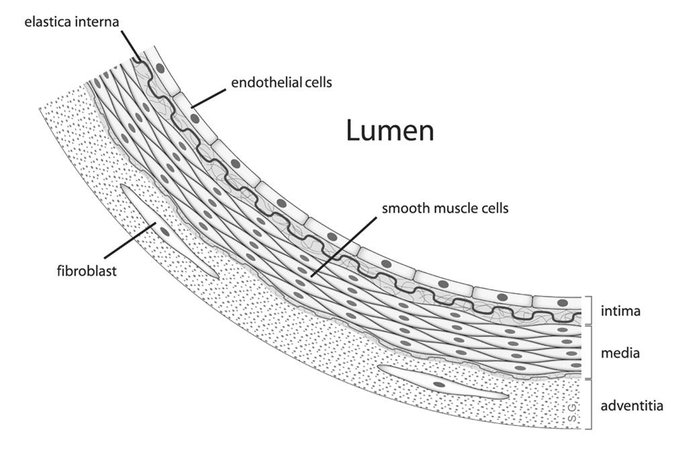
Mes sept années au département de pharmacologie de l’Université de Washington ont été agréables. Oliver (Ollie) Lowry avait été nommé président de ce département environ un an avant mon arrivée. Il était déjà bien reconnu pour ses méthodes ingénieuses impliquant l’enzymologie, la spectrométrie et la fluorométrie dans l’analyse quantitative d’enzymes, de substrats et de produits importants dans des quantités extrêmement faibles de tissus. Il m’a beaucoup aidé à m’initier aux méthodes enzymatiques-spectroscopiques (telles que développées par kalckar) pour l’analyse de l’ATP, de l’ADP et de l’AMP. En tant que nouveau président, Lowry a hérité de deux membres du corps professoral, Helen Graham et Edward Hunter, et en a recruté deux nouveaux, à savoir moi-même et Morris (Morrie) Friedkin. Je n’avais jamais suivi de cours de pharmacologie en tant qu’étudiant, et encore moins enseigné dans un cours, et j’ai donc dû passer beaucoup de temps pendant ma première année à St. Louis devançant les étudiants en médecine. Plus tard, lorsque j’ai créé mon propre département à Brooklyn, j’ai adopté pour le cours de pharmacologie là-bas une grande partie du programme de cours, de laboratoire et de conférence auquel j’avais participé à St. Louis.
Le département de Lowry était un lieu stimulant pour la recherche. Au fil des années où j’étais là-bas, le personnel du ministère a augmenté régulièrement. Lowry a attiré des boursiers postdoctoraux exceptionnels, tels qu’Eli Robbins et Jack Strominger. Nous avons souvent rejoint les membres du département de biochimie de Carl Cori pour des séminaires et des réunions du club de lecture.
Mon premier projet de recherche à l’Université de Washington était une continuation du travail que j’avais commencé à Cornell sur le métabolisme énergétique et la fonction du muscle lisse intestinal du lapin. J’ai pu obtenir une petite bourse pour soutenir mes recherches sur les muscles lisses et embaucher une technicienne, Marilyn (Wales) McCaman, qui est devenue plus tard ma première étudiante diplômée. Vers le milieu de 1951, mon préféré in vitrola préparation des muscles lisses s’était déplacée du duodénum du lapin vers l’aorte thoracique du lapin. J’avais découvert que la bande hélicoïdale (en spirale) de ce vaisseau, correctement coupée et montée dans des chambres d’organes pour l’enregistrement isotonique, donnait des contractions très reproductibles à l’épinéphrine et à la noradrénaline après équilibrage dans une solution de bicarbonate de Krebs oxygénée. J’avais initialement prévu d’étudier les effets des perturbations du métabolisme énergétique sur ces contractions, mais je me suis beaucoup plus intéressé à l’utilisation de la bandelette aortique pour des études sur les interactions médicament-récepteur.
En 1953, j’avais publié un article intitulé « Réactions de bandes d’aorte de lapin à l’épinéphrine, à l’isoprotérénol, au nitrite de sodium et à d’autres médicaments ». Parmi les autres médicaments figurait l’acétylcholine. J’ai constaté qu’il ne produisait que des contractions, qu’il soit ajouté à des bandes de repos ou à des bandes précontractées avec un autre agent. C’était une réponse paradoxale puisque l’acétylcholine était connue pour être un vasodilatateur très puissant in vivo. Je ne soupçonnais pas alors ce que j’ai pu montrer plusieurs années plus tard – à savoir que la relaxation des artères par l’acétylcholine est strictement dépendante de l’endothélium et que ma méthode de préparation des bandelettes a entraîné par inadvertance l’élimination mécanique de toutes les cellules endothéliales.
En 1954, j’ai publié un article sur l’utilisation de la dibénamine dans la différenciation des récepteurs dans la bande aortique, et en 1955 une revue dans Pharmacological Reviews sur la pharmacologie du muscle lisse vasculaire. Dans cette revue, j’ai essayé de développer la théorie des récepteurs comme base logique pour interpréter les réponses du muscle lisse vasculaire à de nombreux neurotransmetteurs, hormones et médicaments. Afin de dériver des équations pour tenir compte de la cinétique d’apparition et de décalage très lente des antagonistes compétitifs par rapport à la cinétique rapide des agonistes, j’ai développé un modèle de biophase dans lequel les agents se déplaçaient entre une phase extracellulaire aqueuse et une phase membranaire lipidique contenant les récepteurs. . Bien que j’aie rendu hommage dans ma critique à AJ Clark pour son travail de pionnier dans le développement de la théorie des récepteurs, J’ai contesté son hypothèse selon laquelle la réponse d’un tissu à un agoniste est proportionnelle à la fraction de récepteurs occupés par l’agoniste. Nos résultats avec la dibénamine, qui s’est comporté comme un inhibiteur compétitif irréversible des adrénergiques ![]() -récepteurs, avait indiqué qu’avec un agoniste puissant comme l’épinéphrine, on pouvait encore atteindre bien plus de la moitié de la contraction maximale lorsque seule une petite fraction des récepteurs était encore active. Ce fut le début de mon intérêt pour le concept de « réserve de récepteurs » ou de « récepteurs de réserve ». (Un an plus tard, RP Stephenson a publié son article classique sur le sujet dans lequel il a introduit les concepts d’efficacité, d’agoniste total et d’agoniste partiel.)
-récepteurs, avait indiqué qu’avec un agoniste puissant comme l’épinéphrine, on pouvait encore atteindre bien plus de la moitié de la contraction maximale lorsque seule une petite fraction des récepteurs était encore active. Ce fut le début de mon intérêt pour le concept de « réserve de récepteurs » ou de « récepteurs de réserve ». (Un an plus tard, RP Stephenson a publié son article classique sur le sujet dans lequel il a introduit les concepts d’efficacité, d’agoniste total et d’agoniste partiel.)
Dans la revue de 1955, j’ai également brièvement rendu compte d’un phénomène nouvellement découvert – à savoir que des bandes d’aorte de lapin subissent une relaxation réversible lorsqu’elles sont exposées à une lumière de longueur d’onde et d’intensité appropriées. Cette photorelaxation était une découverte accidentelle qui provenait de l’observation que, dans une expérience, le tonus contractile actif de deux bandes dans une paire de chambres d’organes fluctuait avec le temps, tandis que celui de deux bandes dans une autre paire de chambres restait stable. Les deux bandes montrant des fluctuations l’ont fait de manière synchrone. Ces deux bandes, mais pas les deux autres, se trouvaient dans des chambres d’orgue près d’une fenêtre à travers laquelle elles étaient exposées à la lucarne. Soupçonnant que les fluctuations de tonalité étaient dues à des fluctuations d’intensité lumineuse sur les bandes près de la fenêtre (c’était une journée nuageuse et lumineuse), J’ai fermé le store de la fenêtre et les deux bandes ont augmenté de ton. J’ai ouvert l’abat-jour et les deux ont diminué de ton. À partir de ce moment, nous n’avons jamais permis que nos bandes soient exposées à la lumière directe du ciel. (Les lampes fluorescentes habituelles au plafond ne produisent pas de photorelaxation.) Certaines études sur les caractéristiques de la photorelaxation ont commencé à Saint-Louis, puis se sont étendues lorsque j’ai déménagé à Brooklyn.
En plus de travailler sur des préparations de muscle lisse in vitro à l’Université de Washington, j’ai également commencé ce qui est devenu de nombreuses années de recherche sur la pharmacologie d’une préparation de muscle cardiaque in vitro – à savoir l’oreillette droite électriquement isolée du cobaye. Pour commencer ce travail, j’ai eu l’aide d’un technicien très compétent, Taisija De Gubareff. En utilisant des méthodes chimiques et enzymatiques pour l’analyse de la créatinine phosphate, de l’ATP, de l’ADP et de l’AMP, nous avons montré que ni le développement d’un « échec expérimental » in vitro(une perte constante de force contractile au fil des heures) ni la récupération après un échec après l’ajout d’un glycoside cardiaque n’étaient dues à des changements de concentration de ces phosphates à haute énergie. Nous avons également rapporté les effets de l’anaérobiose et d’un certain nombre d’agents inotropes positifs et négatifs. Nous avons collaboré avec mon bon ami William Sleator du département de physiologie dans l’étude des changements dans les potentiels d’action cellulaire (mesurés avec des microélectrodes intracellulaires) associés aux changements dans la contractilité de l’oreillette du cobaye en réponse à l’épinéphrine et à l’acétylcholine, et un certain nombre d’autres agents inotropes. Suny Medical Center à Brooklyn (1956-)
Suny Medical Center à Brooklyn (1956-)
En 1956, j’ai accepté le poste de président du nouveau Département de pharmacologie du Collège de médecine de l’Université d’État de New York (SUNY) à New York (en fait à Brooklyn, et plus tard changé de nom en SUNY Downstate Medical Center et plus récemment au SUNY Health Science Center à Brooklyn). Le département faisait auparavant partie d’un département conjoint de physiologie et de pharmacologie dirigé par Chandler Brooks, mais avec l’ouverture d’un nouveau bâtiment de sciences fondamentales relativement énorme (pour l’époque) pour la faculté de médecine et avec un bon soutien financier de l’Université d’État, il était amplement d’espace et de ressources pour un département distinct. De l’ancien département mixte, j’ai hérité Julius Belford comme professeur associé et Bernard Mirkin comme professeur adjoint. Pour des professeurs supplémentaires, j’ai recruté Kwang Soo Lee, Leonard Procita, Lowell Greenbaum, Walter Wosilait et Arthur Zimmerman, tous à temps pour qu’ils donnent notre premier cours aux étudiants en médecine. L’année suivante, CY Kao rejoint le staff. Au cours de la première année également, nous avons accueilli nos premiers étudiants diplômés, à savoir Maurice Feinstein, qui a travaillé avec moi, et Arnold Schwartz, qui a travaillé avec Lee. Au cours de cette année, je n’ai pas fait beaucoup de travail de laboratoire dans le laboratoire de recherche, car la plupart de mon temps a été consacré à l’organisation du département et à l’apprentissage du poste de président. (Je ne suis jamais devenu un administrateur bien organisé et j’ai toujours eu du mal à déléguer des pouvoirs.) et Arnold Schwartz, qui a travaillé avec Lee. Au cours de cette année, je n’ai pas fait beaucoup de travail de laboratoire dans le laboratoire de recherche, car la plupart de mon temps a été consacré à l’organisation du département et à l’apprentissage du poste de président. (Je ne suis jamais devenu un administrateur bien organisé et j’ai toujours eu du mal à déléguer des pouvoirs.) et Arnold Schwartz, qui a travaillé avec Lee. Au cours de cette année, je n’ai pas fait beaucoup de travail de laboratoire dans le laboratoire de recherche, car la plupart de mon temps a été consacré à l’organisation du département et à l’apprentissage du poste de président. (Je ne suis jamais devenu un administrateur bien organisé et j’ai toujours eu du mal à déléguer des pouvoirs.)
Photorelaxation des vaisseaux sanguins
Eugene Greenblatt, mon premier boursier postdoctoral, et Stuart Ehrreich, mon troisième étudiant diplômé, ont participé à cette recherche. Entre autres choses, nous avons pu obtenir un spectre d’action précis (avec un pic à 310 nm) pour la photorelaxation. Plus tard, nous avons observé que l’ajout de nitrite de sodium au milieu de bain sensibilisait fortement la bande aortique de lapin à la photorelaxation et déplaçait le pic du spectre d’action à environ 355 nm. Ehrreich et moi avons découvert que de nombreuses autres préparations de muscles lisses (provenant de l’estomac, de l’intestin et de l’utérus) qui ne se détendaient pas habituellement en réponse aux radiations le faisaient en présence de nitrite inorganique. Percy Lindgren, membre invité du corps professoral de l’ Institut Karolinska, a également travaillé un moment avec nous sur la photosensibilisation par les nitrites.
Mon premier étudiant diplômé à Brooklyn, Maurice Feinstein, a fait son doctorat. recherche de thèse sur les effets de l’insuffisance cardiaque congestive expérimentale, de l’asphyxie et de l’ouabaïne sur les phosphates à haute énergie et la teneur en créatine du cœur du cobaye. Mon deuxième étudiant diplômé, Albert Grossman, qui a commencé à travailler en 1957, a fait sa recherche de thèse sur les effets de la fréquence de stimulation, de la concentration de calcium extracellulaire et de divers médicaments sur l’échange de calcium et la contractilité de l’oreillette gauche du cobaye. Grossman et moi avons publié trois articles basés sur sa recherche de thèse, qui était l’une des premières tentatives pour déterminer les taux d’échange de calcium (en utilisant 45Ca) entre le liquide extracellulaire et divers « pools » intracellulaires de calcium dans le muscle cardiaque dans diverses conditions affectant la contractilité. Nous avons montré que les effets inotropes positifs de la noradrénaline et de la strophanthine-K étaient corrélés à une augmentation du taux d’échange de calcium dans un pool intracellulaire associé au processus contractile et que les effets inotropes négatifs de l’acétylcholine et de l’adénosine étaient corrélés à une diminution du taux d’échange dans ce pool.Nous avons également poursuivi les travaux avec la ryanodine, qui produisait un effet inotrope négatif sur l’oreillette du cobaye et modifiait en fait l’effet force-fréquence d’un escalier positif à négatif (imitant l’escalier normal dans le cœur de grenouille). Sleator, De Gubareff et moi avions montré que la diminution de la force avec la ryanodine (contrairement à celle avec l’acétylcholine ou l’adénosine) n’était pas associée à une diminution de la durée du potentiel d’action. La recherche de thèse de Grossman et quelques années plus tard celle d’un autre étudiant diplômé, Peter Wolf, utilisant également le 45 Ca pour mesurer les effets de la ryanodine sur l’échange de calcium, ont conduit à un modèle hypothétique qui cadre assez bien avec les travaux plus récents d’autres sur les réactions de la ryanodine avec des « récepteurs » impliqués dans le transport du calcium dans le réticulum sarcoplasmique.Mécanismes adrénergiques périphériques
En écrivant la revue de 1955 sur la «Pharmacologie du muscle lisse vasculaire», je m’étais beaucoup intéressé aux mécanismes par lesquels la dénervation postganglionnaire sympathique et certaines drogues comme la cocaïne potentialisent nettement la réponse des organes effecteurs à l’épinéphrine et à la noradrénaline, tout en réduisant nettement la réponse. à la tyramine sympathomimétique. Mon deuxième boursier postdoctoral, Sadashiv (Sada) Kirpekar, a été affecté à ce domaine. Il s’est avéré être un enquêteur doué, et nous avons publié ensemble un certain nombre d’articles sur des travaux menés entre 1959 et 1962. Dans un article, avec pour titre courant « le paradoxe de la cocaïne », nous avons présenté des preuves que dans des bandes aortiques de lapin et des oreillettes électriques isolées du cobaye et du chat, la cocaïne potentialisait les réponses à la noradrénaline et inhibait celles à la tyramine en bloquant un seul et même site sur les terminaisons nerveuses adrénergiques.  Le blocage de ce site a inhibé la captation neuronale de la non adrénaline de la région des récepteurs adrénergiques, potentialisant ainsi son action ; cependant, le blocage du site a également inhibé l’absorption de tyramine, dont l’action sympathomimétique dépend de la libération de noradrénaline à partir des sites de stockage neuronaux, inhibant ainsi son action. Le site, que nous avons appelé le « site de transfert », est devenu plus tard connu sous le nom de site d’absorption-1 (UI). Dans le même article, nous avons montré que la réserpine, qui appauvrissait les granules de stockage neuronal de noradrénaline, n’interférait pas avec l’activité du site d’absorption. En plus de Kirpekar, Peter Cervoni est venu en tant que boursier postdoctoral pour travailler sur les mécanismes adrénergiques périphériques.
Le blocage de ce site a inhibé la captation neuronale de la non adrénaline de la région des récepteurs adrénergiques, potentialisant ainsi son action ; cependant, le blocage du site a également inhibé l’absorption de tyramine, dont l’action sympathomimétique dépend de la libération de noradrénaline à partir des sites de stockage neuronaux, inhibant ainsi son action. Le site, que nous avons appelé le « site de transfert », est devenu plus tard connu sous le nom de site d’absorption-1 (UI). Dans le même article, nous avons montré que la réserpine, qui appauvrissait les granules de stockage neuronal de noradrénaline, n’interférait pas avec l’activité du site d’absorption. En plus de Kirpekar, Peter Cervoni est venu en tant que boursier postdoctoral pour travailler sur les mécanismes adrénergiques périphériques. En 1960, j’ai été invité à présenter un article sur certaines de mes études sur les récepteurs des amines sympathomimétiques lors d’une conférence de la Fondation CIBA sur les mécanismes adrénergiques tenue à la CIBA House à Londres. C’était l’occasion de mon premier voyage à l’étranger et c’était très excitant. Parmi les nombreux pharmacologues distingués présents à la conférence figuraient Sir Henry Dale, Sir John Gaddum et JH Burn. À cette époque, Burn poussait son hypothèse de « lien cholinergique » pour la libération de noradrénaline au niveau des terminaisons nerveuses adrénergiques.
En 1960, j’ai été invité à présenter un article sur certaines de mes études sur les récepteurs des amines sympathomimétiques lors d’une conférence de la Fondation CIBA sur les mécanismes adrénergiques tenue à la CIBA House à Londres. C’était l’occasion de mon premier voyage à l’étranger et c’était très excitant. Parmi les nombreux pharmacologues distingués présents à la conférence figuraient Sir Henry Dale, Sir John Gaddum et JH Burn. À cette époque, Burn poussait son hypothèse de « lien cholinergique » pour la libération de noradrénaline au niveau des terminaisons nerveuses adrénergiques. En 1962-63, j’ai passé une année sabbatique au Département de physiologie de l’Université de Genève, dont Jean Posternak était le directeur. Bien que j’y ai fait des recherches et enseigné, j’ai passé la plupart de mon temps à rédiger des articles sur des recherches que mes collègues et moi avions terminées au cours des années précédentes et sur une revue des mécanismes des récepteurs (voir ci-dessous). J’ai également visité un certain nombre de laboratoires en Europe où des recherches exceptionnelles sur les mécanismes adrénergiques étaient en cours. Parmi ceux-ci figuraient les laboratoires de S. von Euler à Stockholm, E. Muscholl à Mayence et John Gillespie à Glasgow.
En 1962-63, j’ai passé une année sabbatique au Département de physiologie de l’Université de Genève, dont Jean Posternak était le directeur. Bien que j’y ai fait des recherches et enseigné, j’ai passé la plupart de mon temps à rédiger des articles sur des recherches que mes collègues et moi avions terminées au cours des années précédentes et sur une revue des mécanismes des récepteurs (voir ci-dessous). J’ai également visité un certain nombre de laboratoires en Europe où des recherches exceptionnelles sur les mécanismes adrénergiques étaient en cours. Parmi ceux-ci figuraient les laboratoires de S. von Euler à Stockholm, E. Muscholl à Mayence et John Gillespie à Glasgow.
 Au début de 1971, j’ai commencé mon deuxième congé sabbatique, cette fois à la faculté de médecine relativement nouvelle de l’Université de Californie à San Diego (située à La Jolla). Je suis devenu professeur invité à la division de pharmacologie du département de médecine de Steve Mayer. L’une des raisons de ce choix d’un site sabbatique était que je voulais apprendre la méthode d’analyse de l’AMP cyclique que Mayer avait développée (c’était avant le développement des radioimmunodosages pour les nucléotides cycliques). Cependant, je n’ai pas fait beaucoup de recherche à La Jolla, en partie parce qu’une bonne partie de mon temps cette année-là a été consacrée à des fonctions de président de l’American Society for Pharmacology and Experimental Therapeutics.
Au début de 1971, j’ai commencé mon deuxième congé sabbatique, cette fois à la faculté de médecine relativement nouvelle de l’Université de Californie à San Diego (située à La Jolla). Je suis devenu professeur invité à la division de pharmacologie du département de médecine de Steve Mayer. L’une des raisons de ce choix d’un site sabbatique était que je voulais apprendre la méthode d’analyse de l’AMP cyclique que Mayer avait développée (c’était avant le développement des radioimmunodosages pour les nucléotides cycliques). Cependant, je n’ai pas fait beaucoup de recherche à La Jolla, en partie parce qu’une bonne partie de mon temps cette année-là a été consacrée à des fonctions de président de l’American Society for Pharmacology and Experimental Therapeutics.
De retour de La Jolla à Brooklyn en 1972, j’ai poursuivi des recherches sur le rôle des récepteurs situés sur les terminaisons préjonctionnelles (varicosités) des nerfs adrénergiques. J’ai collaboré avec Kirpekar pour tenter de caractériser les  récepteurs inhibiteurs préjonctionnels-adrénergiques sur les terminaisons nerveuses de la rate de chat. Au même moment, un de mes étudiants diplômés, Odd Steinsland, menait un projet de thèse très excitant sur les récepteurs cholinergiques sur les terminaisons nerveuses adrénergiques préjonctionnelles dans l’artère centrale isolée et perfusée du lapin. Il a d’abord caractérisé pharmacologiquement avec l’utilisation de divers agonistes et antagonistes muscariniques le récepteur préjonctionnel à travers lequel l’acétylcholine produit une inhibition marquée de la libération de noradrénaline (contrôlée à la fois par le degré de vasoconstriction et [ 3Libération d’h]norépinéphrine). Il a ensuite étudié la libération de norépinéphrine des neurones adrénergiques de l’artère de l’oreille par des agonistes cholinergiques agissant sur les récepteurs nicotiniques préjonctionnels. Parallèlement, je poursuivais des études, avec l’aide de Taruna Wakade, sur la pharmacologie des récepteurs nicotiniques cholinergiques sur les terminaisons préjonctionnelles adrénergiques de l’oreillette gauche du cobaye.
récepteurs inhibiteurs préjonctionnels-adrénergiques sur les terminaisons nerveuses de la rate de chat. Au même moment, un de mes étudiants diplômés, Odd Steinsland, menait un projet de thèse très excitant sur les récepteurs cholinergiques sur les terminaisons nerveuses adrénergiques préjonctionnelles dans l’artère centrale isolée et perfusée du lapin. Il a d’abord caractérisé pharmacologiquement avec l’utilisation de divers agonistes et antagonistes muscariniques le récepteur préjonctionnel à travers lequel l’acétylcholine produit une inhibition marquée de la libération de noradrénaline (contrôlée à la fois par le degré de vasoconstriction et [ 3Libération d’h]norépinéphrine). Il a ensuite étudié la libération de norépinéphrine des neurones adrénergiques de l’artère de l’oreille par des agonistes cholinergiques agissant sur les récepteurs nicotiniques préjonctionnels. Parallèlement, je poursuivais des études, avec l’aide de Taruna Wakade, sur la pharmacologie des récepteurs nicotiniques cholinergiques sur les terminaisons préjonctionnelles adrénergiques de l’oreillette gauche du cobaye. Théorie et mécanismes des récepteurs
Théorie et mécanismes des récepteurs
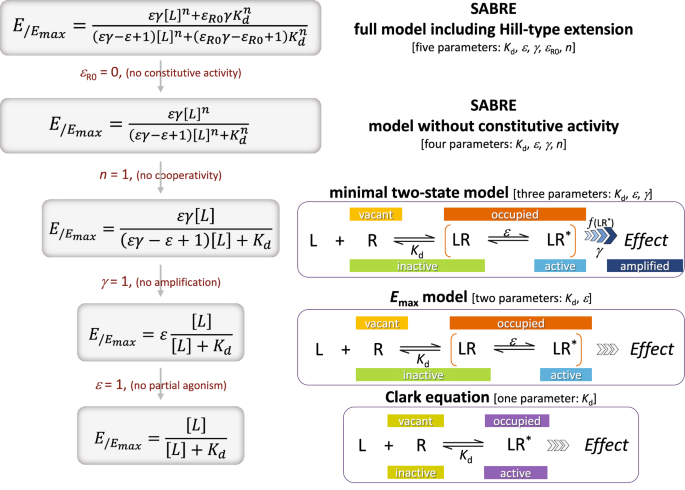
Lorsque j’ai donné pour la première fois un cours sur la théorie et les mécanismes des récepteurs à des étudiants diplômés en 1957-1958, la littérature sur le sujet était relativement rare : des articles de Clark, Gaddum, Schild, Ariëns, Stephenson, Nickerson et moi-même. Je me suis intéressé au développement d’une théorie appropriée (théorie de l’occupation) et de procédures in vitro pour différencier et caractériser les récepteurs. En particulier, je me suis concentré sur les récepteurs des agents adrénergiques et cholinergiques en utilisant comme tissus de test la bande aortique, le segment duodénal et le muscle du fond de l’estomac du lapin, ainsi que l’oreillette gauche et l’anneau trachéal du cobaye à commande électrique.
En 1963, vers la fin de mon année sabbatique à l’Université de Genève, j’ai terminé une revue sur les « Mécanismes des récepteurs » pour le volume 4 de la Revue annuelle de pharmacologie. J’en ai profité pour souligner l’importance des idées de Stephenson sur l’efficacité et les récepteurs de rechange. En 1965, lors d’un symposium sur les mécanismes des récepteurs au Chelsea College de Londres, j’ai présenté un article sur l’utilisation des  -haloalkylamines, comme antagonistes irréversibles des récepteurs, dans la différenciation des récepteurs et dans la détermination des constantes de dissociation des complexes récepteur-agoniste. En utilisant une forme légèrement modifiée des équations de Stephenson et en introduisant un terme,
-haloalkylamines, comme antagonistes irréversibles des récepteurs, dans la différenciation des récepteurs et dans la détermination des constantes de dissociation des complexes récepteur-agoniste. En utilisant une forme légèrement modifiée des équations de Stephenson et en introduisant un terme, pour l’efficacité intrinsèque, j’ai dérivé une équation simple qui prédit que la pente et l’ordonnée à l’origine d’un double tracé réciproque des concentrations équiactives d’un agoniste avant et après l’inactivation irréversible d’une fraction de ses récepteurs, pourraient permettre la détermination à la fois de la fraction de récepteurs encore actifs ainsi que la constante de dissociation (K A ) du complexe agoniste-récepteur. Pour différents agonistes agissant sur le même récepteur, on pourrait calculer à partir du K Avalorise l’occupation fractionnelle par chacun pour obtenir la même réponse standard avant inactivation des récepteurs, et ainsi obtenir des efficacités relatives. En utilisant cette approche, Paula (Bursztyn) Goldberg (une étudiante diplômée) et moi avons comparé les constantes de dissociation et les efficacités relatives des agonistes agissant sur les récepteurs cholinergiques muscariniques de bandes isolées de muscle du fond d’œil de l’estomac de lapin ; et plus tard John Besse (boursier postdoctoral) et moi avons comparé les constantes de dissociation et les efficacités relatives des agonistes agissant sur
pour l’efficacité intrinsèque, j’ai dérivé une équation simple qui prédit que la pente et l’ordonnée à l’origine d’un double tracé réciproque des concentrations équiactives d’un agoniste avant et après l’inactivation irréversible d’une fraction de ses récepteurs, pourraient permettre la détermination à la fois de la fraction de récepteurs encore actifs ainsi que la constante de dissociation (K A ) du complexe agoniste-récepteur. Pour différents agonistes agissant sur le même récepteur, on pourrait calculer à partir du K Avalorise l’occupation fractionnelle par chacun pour obtenir la même réponse standard avant inactivation des récepteurs, et ainsi obtenir des efficacités relatives. En utilisant cette approche, Paula (Bursztyn) Goldberg (une étudiante diplômée) et moi avons comparé les constantes de dissociation et les efficacités relatives des agonistes agissant sur les récepteurs cholinergiques muscariniques de bandes isolées de muscle du fond d’œil de l’estomac de lapin ; et plus tard John Besse (boursier postdoctoral) et moi avons comparé les constantes de dissociation et les efficacités relatives des agonistes agissant sur ![]() 1-récepteurs adrénergiques de l’aorte de lapin. À la lumière de ce que l’on sait maintenant sur les voies de signalisation des récepteurs par les protéines G, il est probablement préférable d’admettre que la procédure pharmacologique que nous avons développée pour obtenir des constantes de dissociation agoniste-récepteur ne peut donner que des valeurs relatives approximatives. Néanmoins, la procédure s’est avérée utile dans un certain nombre d’études.
1-récepteurs adrénergiques de l’aorte de lapin. À la lumière de ce que l’on sait maintenant sur les voies de signalisation des récepteurs par les protéines G, il est probablement préférable d’admettre que la procédure pharmacologique que nous avons développée pour obtenir des constantes de dissociation agoniste-récepteur ne peut donner que des valeurs relatives approximatives. Néanmoins, la procédure s’est avérée utile dans un certain nombre d’études.
 récepteurs bêta-adrénergiques; P. Seeman sur les récepteurs de la dopamine ; S. Snyder et ses collègues sur les récepteurs de la sérotonine et les récepteurs des opiacés). Mon article au symposium était en partie une discussion sur la façon dont les procédures pharmacologiques de différenciation et de caractérisation des récepteurs basées sur la théorie de l’occupation étaient encore très utiles en conjonction avec les nouveaux développements passionnants de la recherche sur les récepteurs réalisés avec des radioligands spécifiques.
récepteurs bêta-adrénergiques; P. Seeman sur les récepteurs de la dopamine ; S. Snyder et ses collègues sur les récepteurs de la sérotonine et les récepteurs des opiacés). Mon article au symposium était en partie une discussion sur la façon dont les procédures pharmacologiques de différenciation et de caractérisation des récepteurs basées sur la théorie de l’occupation étaient encore très utiles en conjonction avec les nouveaux développements passionnants de la recherche sur les récepteurs réalisés avec des radioligands spécifiques.
De plus, j’ai passé en revue les travaux qui avaient été effectués dans mon laboratoire sur les récepteurs bêta-adrénergiques médiant la relaxation du muscle lisse trachéal du cobaye, et j’ai présenté les résultats d’expériences pharmacologiques qui ont montré que ce muscle lisse n’avait pas exclusivement le type 2 du récepteur bêta- adrénergique, selon le dogme de l’époque, mais contenait également un mélange de type bêta1 – généralement sous la forme d’une petite fraction du total des récepteurs bêta, mais, selon le cobaye utilisé, parfois beaucoup plus.
adrénergique, selon le dogme de l’époque, mais contenait également un mélange de type bêta1 – généralement sous la forme d’une petite fraction du total des récepteurs bêta, mais, selon le cobaye utilisé, parfois beaucoup plus.
Endo relaxation dépendante du thélium
Ayant obtenu des preuves pharmacologiques que le muscle lisse trachéal du cobaye possède parfois une fraction importante du récepteur adrénergique de type bêta1 avec le type bêta2 (voir ci-dessus), j’ai décidé qu’il serait bon de réexaminer le muscle lisse de l’aorte thoracique du lapin. pour voir s’il pourrait également avoir des quantités variables du récepteur de type bêta1 mélangé avec le type bêta2. Cependant, dans la toute première expérience conçue pour cette nouvelle étude en mai 1978, une découverte accidentelle à la suite d’une erreur de technicien a complètement changé le cours des recherches dans mon laboratoire. La découverte accidentelle était que sur la préparation d’aorte de lapin utilisée dans l’expérience, les agents muscariniques acétylcholine et carbachol induisaient une relaxation plutôt que la contraction attendue. Pourquoi cette découverte accidentelle était si excitante, comment elle a conduit à notre découverte du facteur relaxant dérivé de l’endothélium (EDRF), et comment ce facteur a finalement été identifié comme étant l’oxyde nitrique ne sera pas discuté ici puisque ces questions seront examinées en détail dans mon Conférence Nobel.![Figure, Physiology of Vasodilation of Vascular Smooth Muscle Image designed by Lana Hariri] - StatPearls - NCBI Bookshelf](https://www.ncbi.nlm.nih.gov/books/NBK556075/bin/Screen__Shot__2019-10-10__at__7.44.43__PM.jpg) En 1982, j’ai démissionné de la présidence du Département de pharmacologie du SUNY Downstate Medical Center, mais j’ai continué en tant que professeur. En 1989, j’ai pris ma retraite de mon poste de professeur (recevant le statut émérite), de sorte que j’étais désormais libre des tâches d’enseignement et des travaux de comité liés au programme de médecine, mais que je pouvais continuer la recherche dans le département. Ma retraite m’a également permis de passer environ trois mois et demi chaque hiver en tant que professeur auxiliaire au Département de pharmacologie moléculaire et cellulaire de l’École de médecine de l’Université de Miami. La plupart de mon temps là-bas, j’ai passé à essayer de rattraper mon retard sur la rédaction de manuscrits et sur la lecture de la littérature en plein essor dans le domaine de la recherche sur l’oxyde nitrique – une tâche impossible de nos jours ! Pendant les séjours d’hiver à Miami, je me tiens au courant de ce qui se passe dans mon laboratoire de recherche à Brooklyn au moyen d’une visite occasionnelle, mais surtout par des communications fréquentes par fax et téléphone avec mon ou mes collègues là-bas. Je me considère très chanceux d’avoir cet arrangement Brooklyn-Miami. Bien sûr, un avantage supplémentaire pour ma femme Maggie et moi est que l’aménagement nous permet de profiter du climat hivernal très agréable à Miami et de certaines des activités de plein air qu’il favorise (le golf, par exemple, dans mon cas).
En 1982, j’ai démissionné de la présidence du Département de pharmacologie du SUNY Downstate Medical Center, mais j’ai continué en tant que professeur. En 1989, j’ai pris ma retraite de mon poste de professeur (recevant le statut émérite), de sorte que j’étais désormais libre des tâches d’enseignement et des travaux de comité liés au programme de médecine, mais que je pouvais continuer la recherche dans le département. Ma retraite m’a également permis de passer environ trois mois et demi chaque hiver en tant que professeur auxiliaire au Département de pharmacologie moléculaire et cellulaire de l’École de médecine de l’Université de Miami. La plupart de mon temps là-bas, j’ai passé à essayer de rattraper mon retard sur la rédaction de manuscrits et sur la lecture de la littérature en plein essor dans le domaine de la recherche sur l’oxyde nitrique – une tâche impossible de nos jours ! Pendant les séjours d’hiver à Miami, je me tiens au courant de ce qui se passe dans mon laboratoire de recherche à Brooklyn au moyen d’une visite occasionnelle, mais surtout par des communications fréquentes par fax et téléphone avec mon ou mes collègues là-bas. Je me considère très chanceux d’avoir cet arrangement Brooklyn-Miami. Bien sûr, un avantage supplémentaire pour ma femme Maggie et moi est que l’aménagement nous permet de profiter du climat hivernal très agréable à Miami et de certaines des activités de plein air qu’il favorise (le golf, par exemple, dans mon cas). De 1982 jusqu’à aujourd’hui, j’ai été récipiendaire de plusieurs distinctions et récompenses pour mes recherches. Naturellement, j’ai été très heureux d’être le récipiendaire. Pourtant, en repensant aux aspects de ma recherche qui m’ont procuré le plus grand plaisir, je ne mettrais pas les honneurs et les récompenses en premier. Je pense que mon plus grand plaisir est venu de chaque première démonstration dans mon laboratoire que des expériences conçues pour tester une nouvelle hypothèse développée pour expliquer une découverte antérieure, souvent déroutante ou paradoxale, ont donné des résultats cohérents avec l’hypothèse. Ce n’est pas seulement le plaisir immédiat d’obtenir de tels résultats, mais aussi le plaisir anticipé de discuter des résultats avec d’autres qui font des recherches dans le même domaine – évidemment un aspect de soutien de l’ego.
De 1982 jusqu’à aujourd’hui, j’ai été récipiendaire de plusieurs distinctions et récompenses pour mes recherches. Naturellement, j’ai été très heureux d’être le récipiendaire. Pourtant, en repensant aux aspects de ma recherche qui m’ont procuré le plus grand plaisir, je ne mettrais pas les honneurs et les récompenses en premier. Je pense que mon plus grand plaisir est venu de chaque première démonstration dans mon laboratoire que des expériences conçues pour tester une nouvelle hypothèse développée pour expliquer une découverte antérieure, souvent déroutante ou paradoxale, ont donné des résultats cohérents avec l’hypothèse. Ce n’est pas seulement le plaisir immédiat d’obtenir de tels résultats, mais aussi le plaisir anticipé de discuter des résultats avec d’autres qui font des recherches dans le même domaine – évidemment un aspect de soutien de l’ego.
Affiliation au moment de l’attribution : SUNY Health Science Center, Brooklyn, NY, États-UnisMotivation du prix : « pour leurs découvertes concernant l’oxyde nitrique en tant que molécule de signalisation dans le système cardiovasculaire »
Sa vie et ses travaux : Robert Furchgott est né à Charleston, en Caroline du Sud, où sa famille dirigeait un grand magasin. Après des études de chimie à Chapel Hill en Caroline du Nord, il obtient son doctorat à la Northwestern University dans l’Illinois. Il a ensuite travaillé à l’Université Cornell à Ithaca, New York, à l’Université Washington à St. Louis et, à partir de 1956, à l’Université d’État de New York à Brooklyn. Furchgott et sa femme Lenore ont eu trois filles. Devenu veuf en 1983, il épouse Margaret Roth, décédée en 2006. Depuis les années 1970, les chercheurs ont compris que le rôle de la couche la plus interne des vaisseaux sanguins, l’endothélium, va au-delà de la protection. En 1980, Robert Furchgott a montré que la capacité des vaisseaux sanguins à se contracter ou à se dilater disparaissait si l’endothélium était retiré. Il a conclu qu’une substance qui provoque l’expansion s’est formée dans cette couche. En 1986, lui et Louis Ignarro, indépendamment l’un de l’autre, ont démontré que cette substance était l’oxyde nitrique (NO). Cette découverte a rendu possibles de nouveaux médicaments, tels que ceux utilisés pour traiter les maladies cardiaques et cardiovasculaires et l’impuissance.
Robert F. Furchgott (1916-2009)Robert Francis Furchgott était un pharmacologue américain qui a partagé (avec Louis J. Ignarro et Ferid Murad) le prix Nobel 1998 de physiologie ou médecine pour la découverte que l’oxyde nitrique (NO) agit comme une molécule de signalisation dans le système cardiovasculaire. Leurs travaux combinés ont découvert un mécanisme entièrement nouveau par lequel les vaisseaux sanguins du corps se détendent et s’élargissent. L’oxyde nitrique (NO), produit par une cellule, agit en pénétrant les membranes et en régulant la fonction d’une autre cellule. Les nerfs et les hormones sont bien connus comme porteurs de signaux, mais cette découverte était un principe de signalisation totalement nouveau dans un système biologique.
https://www.nobelprize.org/prizes/medicine/1998/furchgott/biographical/
https://www.thefamouspeople.com/profiles/robert-f-furchgott-7649.php