
 Premières découvertes d’un fossile de grenouille vieux de 40 millions d’années, trouvé sur l’île Seymour, Antarctique (2015) publié dans «Scientific Reports»
Premières découvertes d’un fossile de grenouille vieux de 40 millions d’années, trouvé sur l’île Seymour, Antarctique (2015) publié dans «Scientific Reports» Première grenouille fossile de l’Antarctique : implications pour les conditions climatiques des hautes latitudes de l’Éocène et le cosmopolitisme gondwanien de l’Australobatrachie
Première grenouille fossile de l’Antarctique : implications pour les conditions climatiques des hautes latitudes de l’Éocène et le cosmopolitisme gondwanien de l’Australobatrachie Les fossiles de grenouilles en Antarctique offrent la preuve d’un passé plus chaud
Les fossiles de grenouilles en Antarctique offrent la preuve d’un passé plus chaud Les fossiles de grenouilles découverts pour la première fois en Antarctique témoignent d’un climat chaud et tempéré dans la péninsule antarctique et de l’existence d’habitats d’eau douce qui abritaient les premiers amphibiens modernes connus il y a 40 millions d’années.
Les fossiles de grenouilles découverts pour la première fois en Antarctique témoignent d’un climat chaud et tempéré dans la péninsule antarctique et de l’existence d’habitats d’eau douce qui abritaient les premiers amphibiens modernes connus il y a 40 millions d’années.
Des fossiles de grenouilles découverts pour la première fois en Antarctique fournissent la preuve d’un climat chaud et tempéré dans la péninsule antarctique et de l’existence d’habitats d’eau douce qui abritaient les premiers amphibiens modernes connus il y a 40 millions d’années, a révélé jeudi une étude publiée dans Scientific Reports .
La découverte d’os fossilisés de la hanche et du crâne vieux de 40 millions d’années de la famille des grenouilles casquées est la première preuve que des amphibiens ou des reptiles à sang froid appartenant à des familles encore existantes se trouvent en Antarctique.
« La grenouille de l’île Seymour rapportée ici est le premier vertébré indicatif d’habitats d’eau douce sur la péninsule antarctique de l’Éocène, après les preuves d’invertébrés et de plantes », rapporte l’étude. Avec la nouvelle découverte, les grenouilles – qui ont la large distribution des amphibiens – ont maintenant été trouvées sur les sept continents. Et les chercheurs émettent maintenant l’hypothèse que « l’Antarctique pourrait avoir agi comme un centre de diversification » pour l’espèce de grenouille casquée, dont des restes ont également été trouvés en Australie.
Avec la nouvelle découverte, les grenouilles – qui ont la large distribution des amphibiens – ont maintenant été trouvées sur les sept continents. Et les chercheurs émettent maintenant l’hypothèse que « l’Antarctique pourrait avoir agi comme un centre de diversification » pour l’espèce de grenouille casquée, dont des restes ont également été trouvés en Australie.
« Jusqu’à présent, l’Antarctique a été considérée comme une voie de dispersion, mais pas comme un lieu d’origine probable. Les nouvelles découvertes de fossiles soutiennent l’hypothèse selon laquelle l’Antarctique pourrait avoir agi comme un centre de diversification pour [les grenouilles casquées] », selon l’étude. Les chercheurs pensent que les fossiles sont la preuve que le climat de la péninsule antarctique à la fin de la période géologique de l’Éocène moyen avant sa séparation du supercontinent sud du Gondwana peut avoir été comparable au climat humide et tempéré des forêts d’Amérique du Sud aujourd’hui, où les cinq vivants les espèces de la grenouille casquée sont exclusivement trouvées.
Les chercheurs pensent que les fossiles sont la preuve que le climat de la péninsule antarctique à la fin de la période géologique de l’Éocène moyen avant sa séparation du supercontinent sud du Gondwana peut avoir été comparable au climat humide et tempéré des forêts d’Amérique du Sud aujourd’hui, où les cinq vivants les espèces de la grenouille casquée sont exclusivement trouvées. Les grenouilles habitent aujourd’hui les basses terres du Chili à l’est des Andes, près des lacs, des étangs et des ruisseaux.
Les grenouilles habitent aujourd’hui les basses terres du Chili à l’est des Andes, près des lacs, des étangs et des ruisseaux.
Les fossiles de grenouilles casquées les plus anciens ont été trouvés en Argentine, selon l’étude.
La découverte suggère que les forêts sud-américaines pourraient être l’équivalent moderne du climat antarctique juste avant la formation des glaciers sur le continent.
Les restes fossiles de grenouille ont été trouvés sur l’île Seymour lors d’expéditions estivales entre 2011 et 2013. Les chercheurs disent que la découverte est un exemple ancien de l’impact du changement climatique sur la survie de différentes espèces, similaire à la façon dont les humains se sont adaptés plus récemment au changement climatique.
Les chercheurs disent que la découverte est un exemple ancien de l’impact du changement climatique sur la survie de différentes espèces, similaire à la façon dont les humains se sont adaptés plus récemment au changement climatique.
« Les découvertes fossiles d’une grenouille et d’un marsupial de l’île Seymour, ainsi que leurs distributions fossiles et récentes, représentent des exemples exceptionnels du rôle du changement climatique mondial sur le changement des aires biogéographiques », a révélé l’étude.
« Malgré le refroidissement global et la disparition des habitats de ces groupes sur de vastes zones allant de l’Antarctique à la Patagonie, ils ont maintenu leur présence relictuelle [de population] dans les forêts de Nothofagus des Andes centrales du Chili », selon l’étude. Le rapport a été publié par Thomas Moers du département de paléobiologie du Musée suédois d’histoire naturelle. Dans un e-mail, Moers a déclaré que la découverte de fossiles de grenouilles casquées est la preuve de l’impact dramatique que le changement climatique peut avoir sur la survie des espèces.
Le rapport a été publié par Thomas Moers du département de paléobiologie du Musée suédois d’histoire naturelle. Dans un e-mail, Moers a déclaré que la découverte de fossiles de grenouilles casquées est la preuve de l’impact dramatique que le changement climatique peut avoir sur la survie des espèces.
« Cela nous dit que des écosystèmes entiers peuvent être anéantis par le changement climatique mondial et que cela pourrait aller vite », a déclaré Moers.
« Même en cas de changement climatique dramatique, certaines espèces (peu nombreuses !) peuvent survivre s’il existe des zones reliques disponibles », a-t-il ajouté. Première grenouille fossile de l’Antarctique : implications pour les conditions climatiques des hautes latitudes de l’Éocène et le cosmopolitisme gondwanien de l’Australobatrachie
Première grenouille fossile de l’Antarctique : implications pour les conditions climatiques des hautes latitudes de l’Éocène et le cosmopolitisme gondwanien de l’Australobatrachie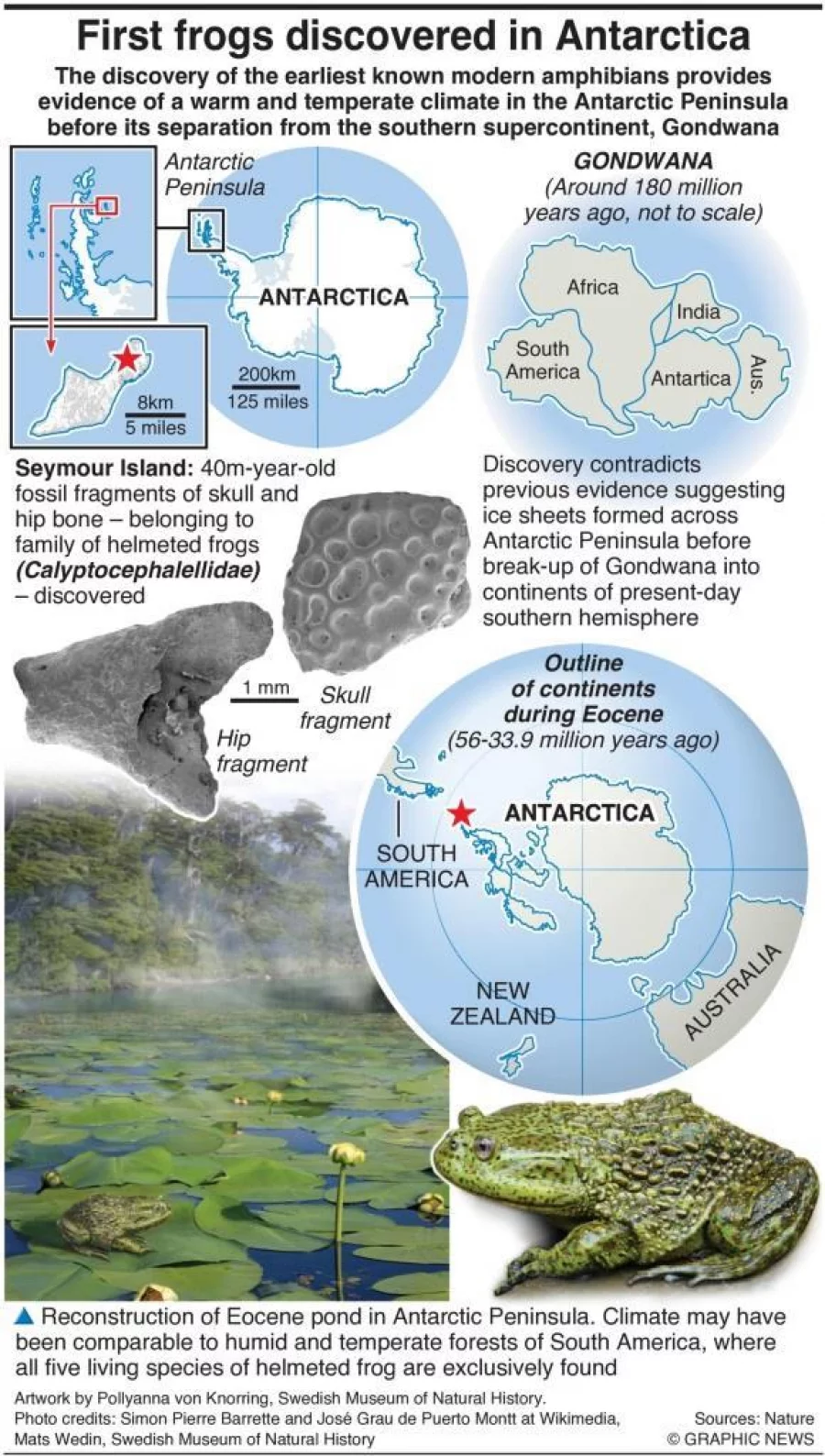 Les tétrapodes continentaux ectothermiques cénozoïques (amphibiens et reptiles) n’ont pas été documentés auparavant de l’Antarctique, contrairement à tous les autres continents. Nous rapportons ici un ilium fossile et un os de crâne orné qui peuvent être attribués à la famille récente, sud-américaine, des anoures Calyptocephalellidae ou grenouilles casquées, représentant le premier amphibien moderne trouvé en Antarctique. Les deux fragments d’os ont été récupérés dans des sédiments de l’Éocène, vieux d’environ 40 millions d’années, sur l’île Seymour, dans la péninsule antarctique. L’enregistrement de grenouilles calyptocéphales hyperossifiées en dehors de l’Amérique du Sud soutient le cosmopolitisme gondwanien du clade anoure Australobatrachia. Nos résultats démontrent que les écosystèmes d’eau douce de l’Éocène en Antarctique ont fourni des habitats favorables aux vertébrés ectothermes (avec des précipitations annuelles moyennes ≥ 900 mm,
Les tétrapodes continentaux ectothermiques cénozoïques (amphibiens et reptiles) n’ont pas été documentés auparavant de l’Antarctique, contrairement à tous les autres continents. Nous rapportons ici un ilium fossile et un os de crâne orné qui peuvent être attribués à la famille récente, sud-américaine, des anoures Calyptocephalellidae ou grenouilles casquées, représentant le premier amphibien moderne trouvé en Antarctique. Les deux fragments d’os ont été récupérés dans des sédiments de l’Éocène, vieux d’environ 40 millions d’années, sur l’île Seymour, dans la péninsule antarctique. L’enregistrement de grenouilles calyptocéphales hyperossifiées en dehors de l’Amérique du Sud soutient le cosmopolitisme gondwanien du clade anoure Australobatrachia. Nos résultats démontrent que les écosystèmes d’eau douce de l’Éocène en Antarctique ont fourni des habitats favorables aux vertébrés ectothermes (avec des précipitations annuelles moyennes ≥ 900 mm, Introduction : Conformément aux preuves géologiques, il a été émis l’hypothèse que la formation des calottes glaciaires de l’Antarctique est antérieure à la rupture finale du Gondwana, à l’ouverture du passage de Drake et à l’isolement thermique du continent. Cela se reflète par une faible diversité de mammifères terrestres sur la péninsule antarctique au cours de l’Éocène moyen à tardif avec seulement deux espèces de grands mammifères et dix espèces de petits mammifères qui contraste fortement avec la faune marine très diversifiée indiquant une faune tempérée. conditions dans la mer de Weddell . Cependant, aucun vertébré continental ectothermique cénozoïque (poissons d’eau douce, amphibiens et reptiles) n’a été connu jusqu’à présent en Antarctique. Nous rapportons ici la découverte d’un ilium fossile de l’île Seymour, péninsule antarctique qui peut être attribué à l’ordre des lissamphibiens Anura, et un fragment d’os crânien sculpté qui dérive très probablement d’un anoure hyperossifié. Nous attribuons les spécimens au genre sud-américain Calyptocephalella . Les Calyptocephalellids, ou grenouilles casquées, sont largement connues en Patagonie depuis le Crétacé supérieur. Ils se sont éteints en Patagonie argentine au Miocène, probablement lié à une diminution de l’humidité causée par la montée des Andes, puisque la famille a survécu jusqu’à nos jours dans un refuge tempéré mais humide des Andes centrales chiliennes .
Introduction : Conformément aux preuves géologiques, il a été émis l’hypothèse que la formation des calottes glaciaires de l’Antarctique est antérieure à la rupture finale du Gondwana, à l’ouverture du passage de Drake et à l’isolement thermique du continent. Cela se reflète par une faible diversité de mammifères terrestres sur la péninsule antarctique au cours de l’Éocène moyen à tardif avec seulement deux espèces de grands mammifères et dix espèces de petits mammifères qui contraste fortement avec la faune marine très diversifiée indiquant une faune tempérée. conditions dans la mer de Weddell . Cependant, aucun vertébré continental ectothermique cénozoïque (poissons d’eau douce, amphibiens et reptiles) n’a été connu jusqu’à présent en Antarctique. Nous rapportons ici la découverte d’un ilium fossile de l’île Seymour, péninsule antarctique qui peut être attribué à l’ordre des lissamphibiens Anura, et un fragment d’os crânien sculpté qui dérive très probablement d’un anoure hyperossifié. Nous attribuons les spécimens au genre sud-américain Calyptocephalella . Les Calyptocephalellids, ou grenouilles casquées, sont largement connues en Patagonie depuis le Crétacé supérieur. Ils se sont éteints en Patagonie argentine au Miocène, probablement lié à une diminution de l’humidité causée par la montée des Andes, puisque la famille a survécu jusqu’à nos jours dans un refuge tempéré mais humide des Andes centrales chiliennes .Cartes et colonne stratigraphique de la zone étudiée et succession. (a) Carte de l’Antarctique montrant l’emplacement de la zone d’étude. (b) Carte du nord de la péninsule antarctique indiquant l’emplacement de l’île Seymour. (c) Esquisse géologique de l’île Seymour, montrant la position de la localité fossile (astérisque). (d) Colonne stratigraphique de la Formation de La Meseta sur l’île Seymour (de ) indiquant la position de la localité fossile (astérisque). Redessiné à partir de .
Le matériel décrit ici provient de dépôts estuariens à marins marginaux de la formation Eocène La Meseta qui ont été déposés dans le bassin de James Ross, un bassin d’arrière-arc à l’est de la péninsule antarctique, et qui sont largement exposés sur l’île Seymour ( Fig. 1b, c ). La localité fossilifère IAA 2/95, également connue sous le nom de « site marsupial », est une lentille de quelques m 2 de large constituée d’un conglomérat coquillier mal consolidé. Il est situé dans la partie centrale de l’ Allomember Cucullaea I, au sein de l’unité Telm 5 sur le versant nord-ouest de la mesa (Fig. 1c,d ), et officieusement appelé « horizon Natica ». Il a produit des dents de requin, de raie et de raie, des restes de poissons marins osseux, ainsi que des dents de mammifères terrestres, des cocons de vers (clitellés) et des graines de nénuphars. Sur la base des occurrences de dinokystes, l’âge de ce gisement est estimé à environ 40 Ma (Bartonien, Éocène) .
Les restes fossiles de grenouilles ont été collectés au cours de trois expéditions conjointes argentino-suédoises sur l’île Seymour au cours des étés australs 2011–13. Les fragments d’os ont été concentrés à partir d’échantillons de sédiments tamisés à sec comme décrit par et triés à l’aide d’un stéréomicroscope Leica MZ6. Le matériel est conservé dans les collections paléozoologiques du Musée suédois d’histoire naturelle de Stockholm, avec les numéros d’inventaire NRM-PZ B281 et B282.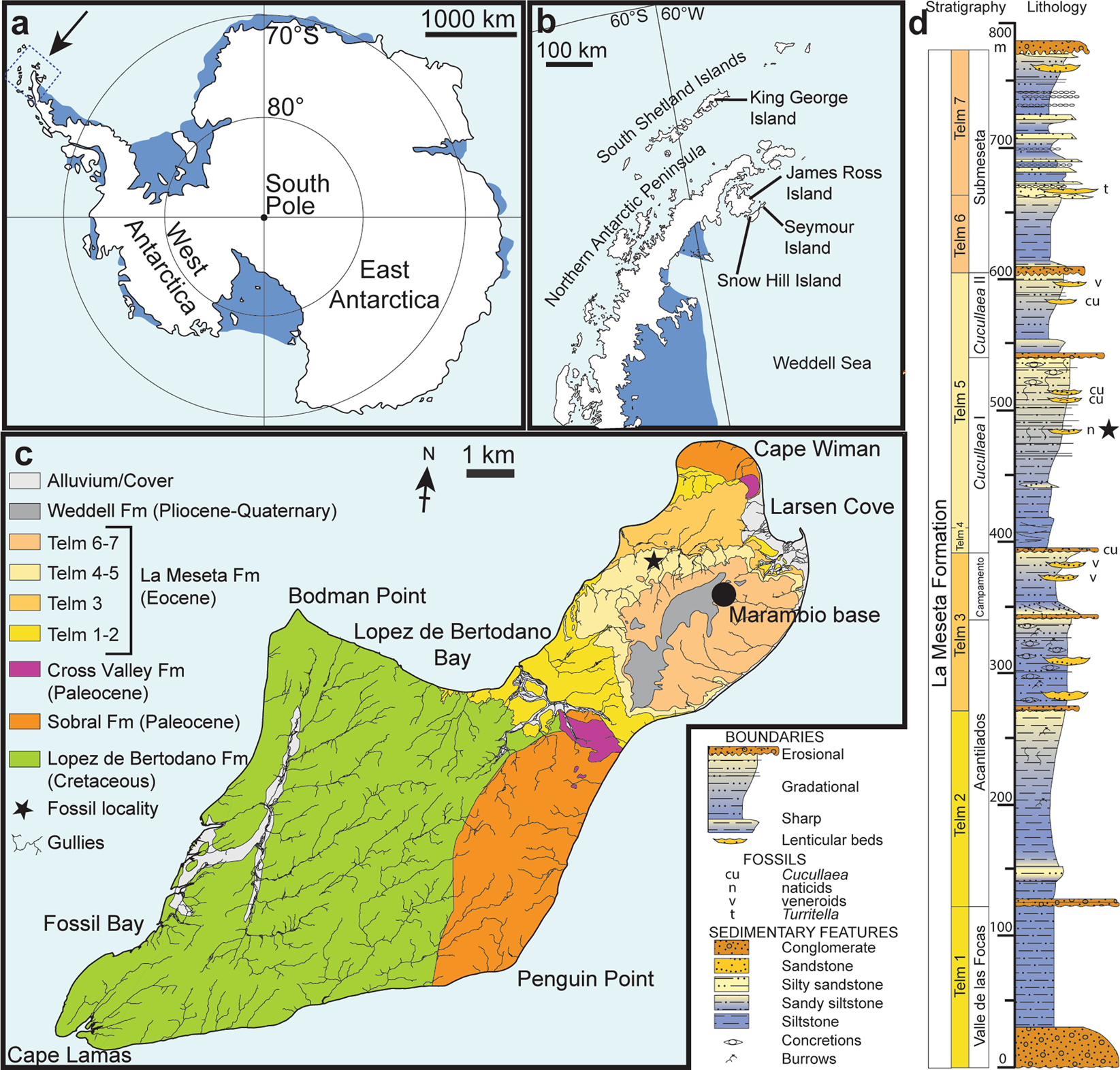 Résultats – Paléontologie systématique
Résultats – Paléontologie systématique
Anura Fischer von Waldheim, 1813 – Neobatrachia Reig, 1958 Australobatrachie Frost et al ., 2006 – Calyptocephalellidae Reig, 1960 Calyptocephalella Strand, 1928 –Calyptocephalella sp. Figures 2 et 3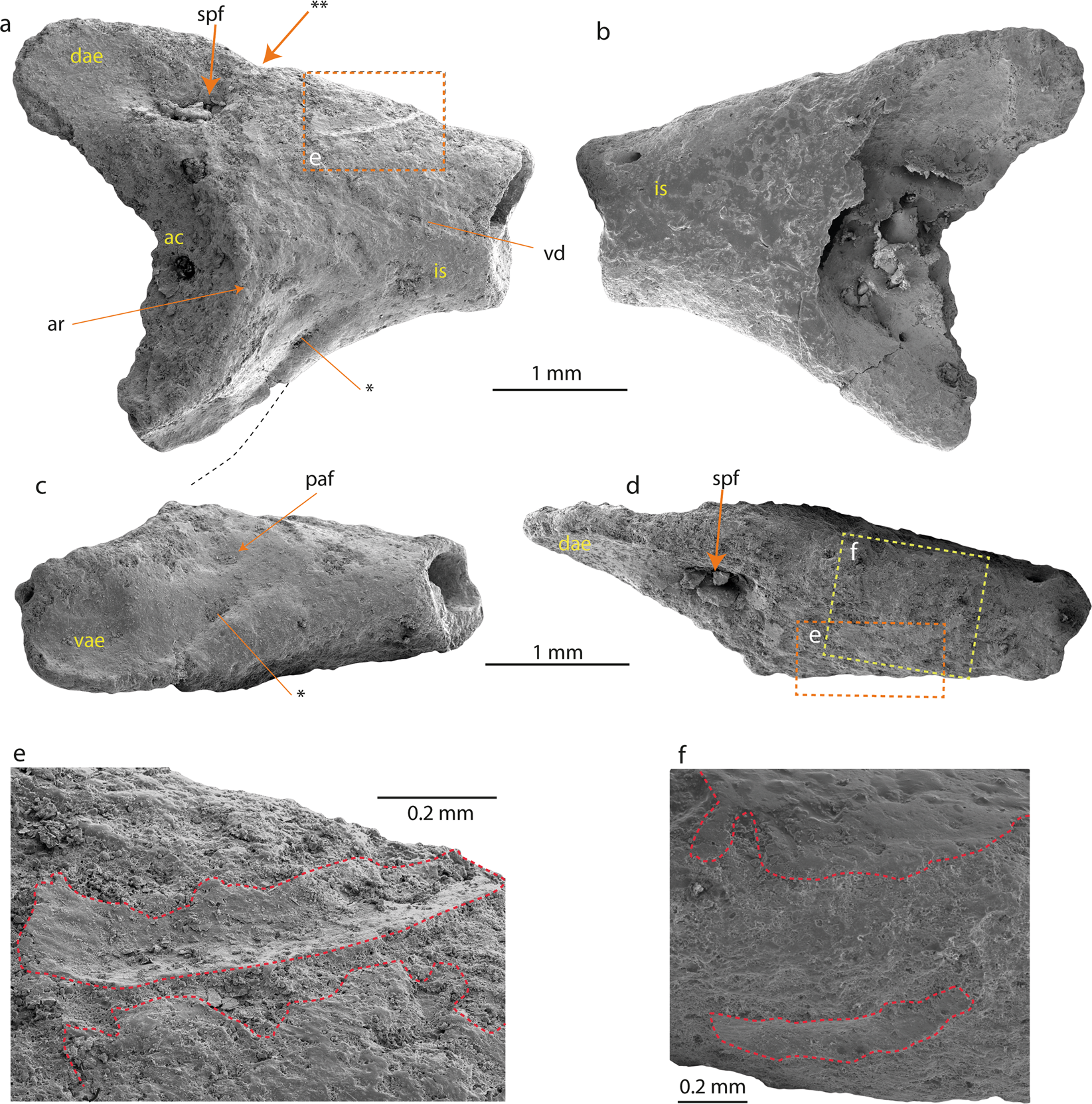 Ilium (NRM-PZ B282) de Calyptocephalella sp. de l’île Seymour, Antarctique. Ilium en vues latérale ( a ), médiale ( b ), ventrale ( c ) et dorsale ( d ). Région agrandie de la protubérance dorsale en vue latérale ( e – rectangle en pointillés de couleur orange) et dorsale ( f – rectangle en pointillés de couleur jaune). La ligne pointillée en noir sur ( a ) indique le contour probable de l’extension postérieure de l’expansion acétabulaire ventrale. L’astérisque (*) sur ( a , c) indique la dépression peu profonde et large de l’expansion acétabulaire ventrale. Le double astérisque (**) indique l’encoche caudalement à partir de la protubérance dorsale. Les lignes rouges en pointillés sur ( e , f ) décrivent la surface osseuse intacte. Abréviations : ac, acétabulum ; ar, jante acétabulaire ; dae, expansion acétabulaire dorsale ; est, arbre iliaque ; paf, fosse préacétabulaire ; spf, fosse supraacétabulaire ; vae, expansion acétabulaire ventrale ; vd, dépression ventrale.
Ilium (NRM-PZ B282) de Calyptocephalella sp. de l’île Seymour, Antarctique. Ilium en vues latérale ( a ), médiale ( b ), ventrale ( c ) et dorsale ( d ). Région agrandie de la protubérance dorsale en vue latérale ( e – rectangle en pointillés de couleur orange) et dorsale ( f – rectangle en pointillés de couleur jaune). La ligne pointillée en noir sur ( a ) indique le contour probable de l’extension postérieure de l’expansion acétabulaire ventrale. L’astérisque (*) sur ( a , c) indique la dépression peu profonde et large de l’expansion acétabulaire ventrale. Le double astérisque (**) indique l’encoche caudalement à partir de la protubérance dorsale. Les lignes rouges en pointillés sur ( e , f ) décrivent la surface osseuse intacte. Abréviations : ac, acétabulum ; ar, jante acétabulaire ; dae, expansion acétabulaire dorsale ; est, arbre iliaque ; paf, fosse préacétabulaire ; spf, fosse supraacétabulaire ; vae, expansion acétabulaire ventrale ; vd, dépression ventrale.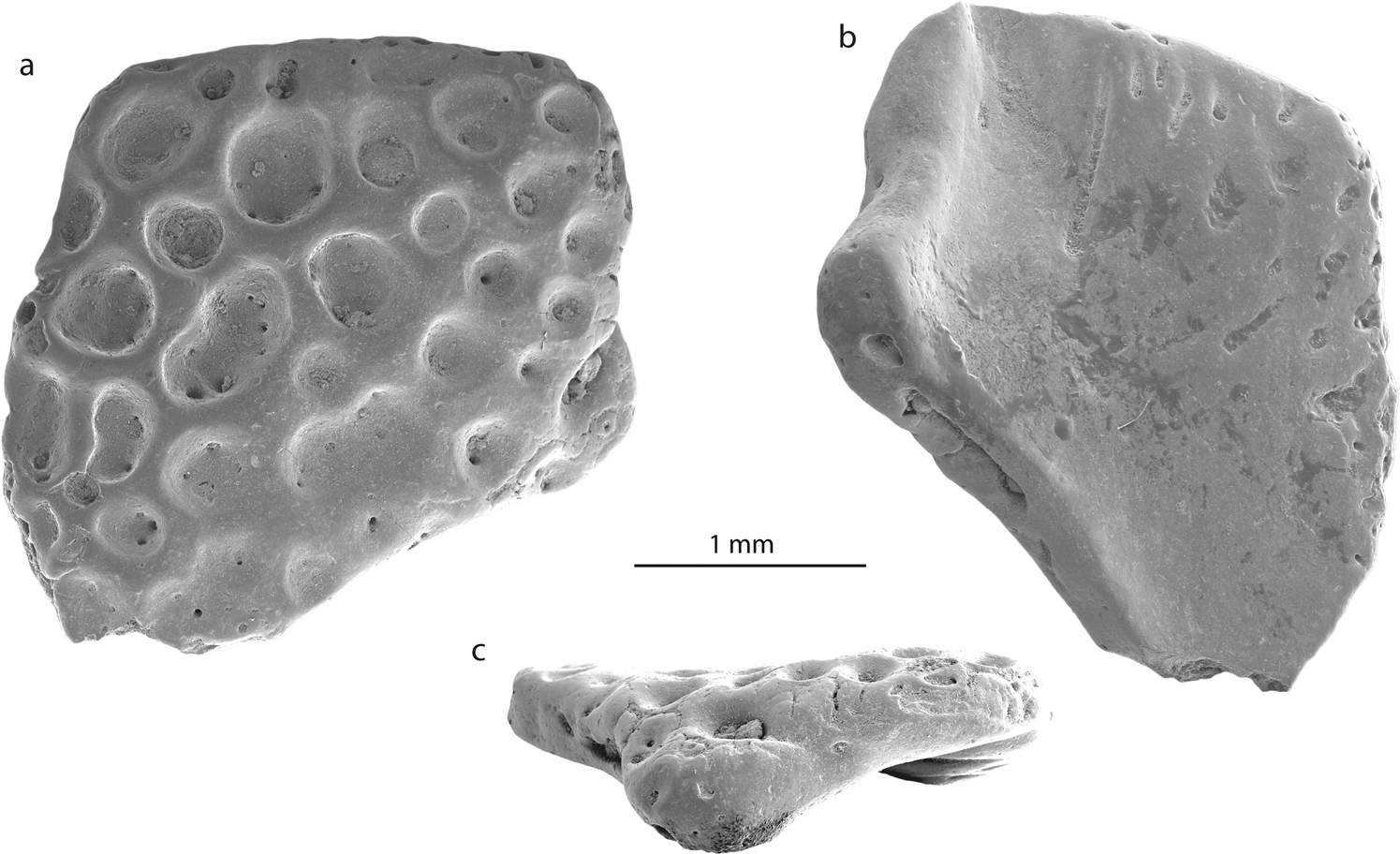 Fragment d’os crânien (NRM-PZ B281) de Calyptocephalella sp. de l’île Seymour, Antarctique en vues dorsale (a), ventrale ( b) et latérale (c).
Fragment d’os crânien (NRM-PZ B281) de Calyptocephalella sp. de l’île Seymour, Antarctique en vues dorsale (a), ventrale ( b) et latérale (c).
Spécimens référés : Musée suédois d’histoire naturelle NRM-PZ B282, ilium droit (Fig. 2 ), NRM-PZ B281, os du crâne (Fig. 3 ).
Localité, horizon et âge : IAA 2/95, site marsupial, île Seymour, péninsule Antarctique (64°13′58″S ; 56°39′06″O). ‘Horizon Natica’ dans l’Allomembre Cucullaea I (Telm 5) de la Formation de La Meseta, Bartonien (40 Ma), Éocène (Fig. 1 ).
Des mesures : La partie préservée de l’ilium mesure 3,9 mm de longueur, la distance entre la pointe de l’expansion acétabulaire dorsale et la pointe (conservée) de l’expansion acétabulaire ventrale mesure 3,3 mm, la hauteur la plus élevée de la fosse acétabulaire est égale à 2,5 mm. L’os du crâne mesure 2,7 mm dans ses parties les plus larges et les plus longues. Ilion : L’ilium droit fragmentaire (NRM-PZ B282) est dépourvu de la partie caudale de l’acétabulum et de la majeure partie de la diaphyse iliaque. L’expansion acétabulaire dorsale a une surface latérale lisse et est plus haute que la partie conservée de l’expansion acétabulaire ventrale (Fig. 2a ). Une fosse supraacétabulaire large et profonde est présente à sa base (Fig. 2a,d ). La partie préservée de l’acétabulum est concave et sa forme permet de conclure un contour (semi-)circulaire. Le bord acétabulaire est le plus proéminent à sa partie antérieure (Fig. 2a,c). L’expansion acétabulaire ventrale à peine développée se projette ventralement. La partie la plus postérieure de l’expansion acétabulaire ventrale est rompue. Cependant, la partie antérieure de l’expansion acétabulaire ventrale est plus haute que la partie postérieure conservée. En vue ventrale (Fig. 2a,c ), la surface latérale de l’expansion acétabulaire ventrale est convexe. L’expansion acétabulaire ventrale possède une dépression peu profonde et large.
Ilion : L’ilium droit fragmentaire (NRM-PZ B282) est dépourvu de la partie caudale de l’acétabulum et de la majeure partie de la diaphyse iliaque. L’expansion acétabulaire dorsale a une surface latérale lisse et est plus haute que la partie conservée de l’expansion acétabulaire ventrale (Fig. 2a ). Une fosse supraacétabulaire large et profonde est présente à sa base (Fig. 2a,d ). La partie préservée de l’acétabulum est concave et sa forme permet de conclure un contour (semi-)circulaire. Le bord acétabulaire est le plus proéminent à sa partie antérieure (Fig. 2a,c). L’expansion acétabulaire ventrale à peine développée se projette ventralement. La partie la plus postérieure de l’expansion acétabulaire ventrale est rompue. Cependant, la partie antérieure de l’expansion acétabulaire ventrale est plus haute que la partie postérieure conservée. En vue ventrale (Fig. 2a,c ), la surface latérale de l’expansion acétabulaire ventrale est convexe. L’expansion acétabulaire ventrale possède une dépression peu profonde et large.  Dans la zone préacétabulaire, une petite fosse préacétabulaire peu profonde est présente (Fig. 2c). La partie préservée de la diaphyse iliaque est endommagée et empêche de dire avec certitude si la protubérance dorsale est présente ou absente. Un sillon longitudinal étroit et peu profond est observable dans la face latérale de la diaphyse iliaque, qui correspond probablement au prolongement postérieur de la dépression ventrale (sensu 10 ) (Fig. 2a ). Cependant, des parties intactes de la surface osseuse sont conservées légèrement ventrales à la marge dorsale sur les surfaces latérale et médiale (Fig. 2e, f ).
Dans la zone préacétabulaire, une petite fosse préacétabulaire peu profonde est présente (Fig. 2c). La partie préservée de la diaphyse iliaque est endommagée et empêche de dire avec certitude si la protubérance dorsale est présente ou absente. Un sillon longitudinal étroit et peu profond est observable dans la face latérale de la diaphyse iliaque, qui correspond probablement au prolongement postérieur de la dépression ventrale (sensu 10 ) (Fig. 2a ). Cependant, des parties intactes de la surface osseuse sont conservées légèrement ventrales à la marge dorsale sur les surfaces latérale et médiale (Fig. 2e, f ). 
L’ilium droit fragmentaire peut être référé à un anoure sur la base des caractères suivants (les nombres avant les caractères correspondent aux numéros des caractéristiques de l’annexe 1 dans Gardner et al . ) : 7 . cotyle (semi-) circulaire ; 9 . acétabulum à marges distinctes ; 10 . surface acétabulaire concave ; 13 . au moins l’expansion acétabulaire dorsale est fortement divergente ; 18 . la protubérance dorsale présente. Ainsi, l’ilium dérive d’une grenouille de petite taille (3,8 ± 0,4 cm de longueur museau-évent, voir méthodes, tableau 1). Le spécimen est en partie érodé et plutôt mal conservé ; cependant, il peut être comparé à toutes les familles de grenouilles sud-américaines et australiennes (Figs. , S1 et S2 , Tableau S1 ). Les familles Ranidae, Bufonidae et Hylidae n’ont pas été illustrées dans le présent travail, car leur morphologie est bien connue (Tableau S1 ). La comparaison a été faite au niveau familial, puisque l’ilia présente des traits diagnostiques caractéristiques d’identification de la famille (dimensions des expansions acétabulaires dorsale et ventrale ; localisation de la protubérance dorsale par rapport au bord antérieur du rebord acétabulaire etc.).  L’ilium étudié (NRM-PZ B282) diffère par : (1) la partie antérieure réduite de l’expansion acétabulaire dorsale de presque toutes les familles de grenouilles sud-américaines et australiennes et du genre Telmatobufo, qui ont une partie antérieure modérément ou fortement développée de l’expansion acétabulaire dorsale . Seuls le genre Calyptocephalella (Fig. 4c,e ), les familles Ranidae , Pipidae (Fig. S1a ), Rhinodermatidae (Fig. S1j ) et Leptodactylidae (Fig. S2b) ont un état/morphologie similaire de ce personnage. (2) Protubérance dorsale située au niveau ou en avant du bord antérieur du rebord acétabulaire de presque toutes les familles, à l’exception des Brachycephalidae (Fig. S1a ), des Rhinodermatidae (Fig. S1j ), des Telmatobiidae (Fig. S1k ), des Hyloididae ( Fig. S1m ), Leptodactylidae (Fig. S2b ) et les genres Calyptocephalella (Fig. 4b–e ) et Telmatobufo (Fig. 4g, h ). (3) Développement de l’expansion acétabulaire dorsale des familles Ranidae, Hylidae, Bufonidae, Myobatrachidae (Fig. 4i ), Pipidae (Fig. S1i ), Microhylidae (Fig. S1b ), Telmatobiidae (Fig. S1k ), Leptodactylidae (Fig. S2b ), Allophrynidae (Fig. S2c ), Centrolenidae (Fig. S2d ) et le genre Telmatobufo (Fig. .4g ,h ). D’autres familles ont une expansion acétabulaire dorsale modérément ou bien développée, cependant, en raison d’une conservation incomplète de la grenouille antarctique, toute autre comparaison est impossible. (4) Protubérance dorsale faiblement développée et absence de tubercule dorsal de presque toutes les familles (par exemple Limnodynastidae, Fig. 4j ), en plus des Calyptocephalellidae (Fig. 4b–e,g,h ), des Myobatrachidae (Fig. 4i ), Craugastoridae (Fig. S1e ) et Dendrabatidae (Fig. S2e ).
L’ilium étudié (NRM-PZ B282) diffère par : (1) la partie antérieure réduite de l’expansion acétabulaire dorsale de presque toutes les familles de grenouilles sud-américaines et australiennes et du genre Telmatobufo, qui ont une partie antérieure modérément ou fortement développée de l’expansion acétabulaire dorsale . Seuls le genre Calyptocephalella (Fig. 4c,e ), les familles Ranidae , Pipidae (Fig. S1a ), Rhinodermatidae (Fig. S1j ) et Leptodactylidae (Fig. S2b) ont un état/morphologie similaire de ce personnage. (2) Protubérance dorsale située au niveau ou en avant du bord antérieur du rebord acétabulaire de presque toutes les familles, à l’exception des Brachycephalidae (Fig. S1a ), des Rhinodermatidae (Fig. S1j ), des Telmatobiidae (Fig. S1k ), des Hyloididae ( Fig. S1m ), Leptodactylidae (Fig. S2b ) et les genres Calyptocephalella (Fig. 4b–e ) et Telmatobufo (Fig. 4g, h ). (3) Développement de l’expansion acétabulaire dorsale des familles Ranidae, Hylidae, Bufonidae, Myobatrachidae (Fig. 4i ), Pipidae (Fig. S1i ), Microhylidae (Fig. S1b ), Telmatobiidae (Fig. S1k ), Leptodactylidae (Fig. S2b ), Allophrynidae (Fig. S2c ), Centrolenidae (Fig. S2d ) et le genre Telmatobufo (Fig. .4g ,h ). D’autres familles ont une expansion acétabulaire dorsale modérément ou bien développée, cependant, en raison d’une conservation incomplète de la grenouille antarctique, toute autre comparaison est impossible. (4) Protubérance dorsale faiblement développée et absence de tubercule dorsal de presque toutes les familles (par exemple Limnodynastidae, Fig. 4j ), en plus des Calyptocephalellidae (Fig. 4b–e,g,h ), des Myobatrachidae (Fig. 4i ), Craugastoridae (Fig. S1e ) et Dendrabatidae (Fig. S2e ).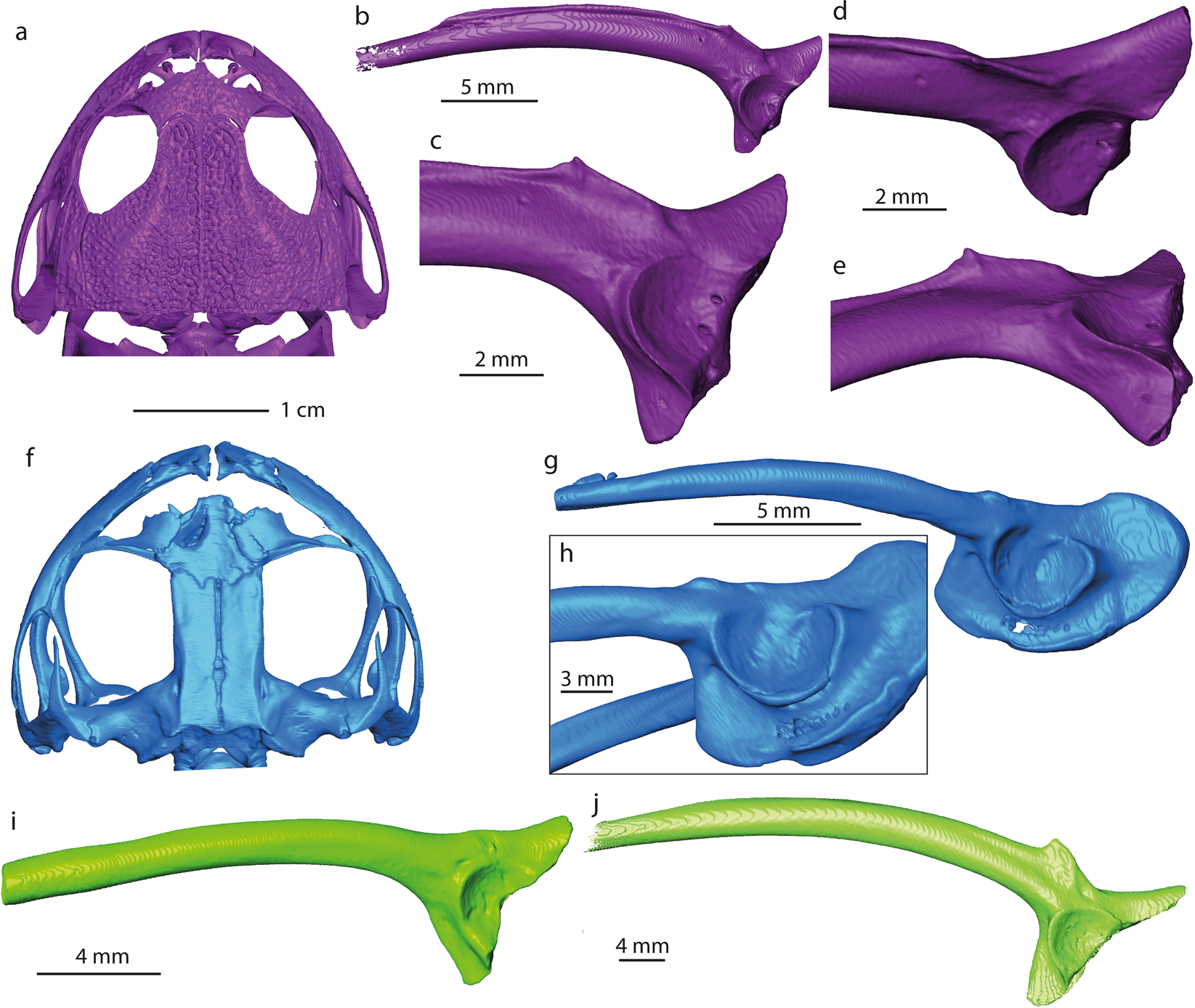 Modèles 3D de certains éléments squelettiques d’Australobatrachie. ( a,f ) crâne et ( b – e , g – j ) ilia de Calyptocephalella gayi ( a–e ); Telmatobufo venustus ( f – h ); Myobatrachus gouldii ( i ); Limnodynastes convexiusculus ( j ). Les numéros de collecte de chaque spécimen sont répertoriés dans le tableau S1
Modèles 3D de certains éléments squelettiques d’Australobatrachie. ( a,f ) crâne et ( b – e , g – j ) ilia de Calyptocephalella gayi ( a–e ); Telmatobufo venustus ( f – h ); Myobatrachus gouldii ( i ); Limnodynastes convexiusculus ( j ). Les numéros de collecte de chaque spécimen sont répertoriés dans le tableau S1
Parmi les formes comparées, seul le genre endémique sud-américain Calyptocephalella ressemble aux quatre caractères mentionnés. En plus de cela, une dépression peu profonde et large sur la partie antérieure de l’expansion acétabulaire ventrale est un caractère unique observable sur notre ilion (NRM-PZ B282) et Calyptocephalella gayi récente (Fig. 4 ). De plus, l’ilium fossile présente une dépression ventrale sur sa surface latérale en avant de l’acétabulum (Fig. 2c ), une structure comparable peut également être observée chez l’espèce fossile Calyptocephalella canqueli mais pas chez l’espèce récente C. gayi (Fig. 4 ).
Élément de crâne : Le deuxième fragment osseux (NRM-PZ B281) est plat et légèrement incurvé. Les deux côtés de l’os ont des structures différentes. Une surface est recouverte de petites à grandes cavités circulaires ou réniformes, plutôt profondes, qui s’enfoncent dans la surface plane de l’os (Fig. 3a ). Les diamètres des fosses varient de 0,1 à 0,7 mm et certaines d’entre elles sont perforées par des foramina. La face opposée de l’os est en général lisse, légèrement creusée et percée de quelques foramens dont certains sont précédés d’une rainure (Fig.3b ). Un côté du fragment préserve une marge ininterrompue de l’os d’origine avec un processus distinct qui est plié et qui donne à l’os une forme incurvée (Fig. 3c). La surface ornée de l’os se projette légèrement au-dessus de ce processus.  Une ornementation comparable, constituée de fosses de tailles différentes, se trouve sur les surfaces dorsales de différents os crâniens et post-crâniens d’amphibiens et de reptiles. Parmi eux, les groupes suivants peuvent être exclus : (1) Albanerpetontidae (Allocaudata) ; Les albanerpetontidés sont un groupe primaire de lissamphibiens Laurasiens avec une seule occurrence en Afrique du Nord. Jusqu’à présent, aucune preuve d’un rayonnement gondwanien des albanerpétontides n’existe. En plus de cela, tous leurs os ornementés (par exemple frontaux, prémaxillaires) ne ressemblent pas à l’os décrit ici. (2) Caudata ; les salamandres sont également considérées comme un groupe laurasien, avec un certain nombre d’occurrences en Afrique qui nécessitent une révision critique . Chez les salamandres, les os ornés se trouvent à la fois parmi les os du crâne et sur les vertèbres (sur des plaques situées à l’extrémité de l’arc neural) . Ornementation osseuse ici (par exemple Tylototriton, Chelotriton, Echinotriton ) est représenté par un réseau de fosses, des crêtes et des épines pointues qui ne ressemblent pas à l’os décrit ici.
Une ornementation comparable, constituée de fosses de tailles différentes, se trouve sur les surfaces dorsales de différents os crâniens et post-crâniens d’amphibiens et de reptiles. Parmi eux, les groupes suivants peuvent être exclus : (1) Albanerpetontidae (Allocaudata) ; Les albanerpetontidés sont un groupe primaire de lissamphibiens Laurasiens avec une seule occurrence en Afrique du Nord. Jusqu’à présent, aucune preuve d’un rayonnement gondwanien des albanerpétontides n’existe. En plus de cela, tous leurs os ornementés (par exemple frontaux, prémaxillaires) ne ressemblent pas à l’os décrit ici. (2) Caudata ; les salamandres sont également considérées comme un groupe laurasien, avec un certain nombre d’occurrences en Afrique qui nécessitent une révision critique . Chez les salamandres, les os ornés se trouvent à la fois parmi les os du crâne et sur les vertèbres (sur des plaques situées à l’extrémité de l’arc neural) . Ornementation osseuse ici (par exemple Tylototriton, Chelotriton, Echinotriton ) est représenté par un réseau de fosses, des crêtes et des épines pointues qui ne ressemblent pas à l’os décrit ici.  (3) Crocodylie ; chez les crocodyliformes, des modèles d’ornementation comparables avec des fosses bien développées n’apparaissent qu’avec la croissance au cours des stades ontogénétiques ultérieurs .
(3) Crocodylie ; chez les crocodyliformes, des modèles d’ornementation comparables avec des fosses bien développées n’apparaissent qu’avec la croissance au cours des stades ontogénétiques ultérieurs . 
Le motif d’ornementation trouvé dans NRM-PZ B281 est comparable à celui de certains genres de grenouilles, c’est-à-dire Thaumastosaurus , Beelzebufo , Calyptocephalella et Baurubatrachus , mais seuls les trois derniers genres sont des formes gondwaniennes et, par conséquent, considérés pour comparaison ici. Beelzebufo est une forme très grande et le motif d’ornementation est présent à la fois sur les os du crâne et sur les vertèbres. Calyptocephalella et Baurubatrachus ont des motifs d’ornementation très similaires sur les surfaces des os du crâne hyperossifiés, comparables à notre spécimen.  Une analyse phylogénétique récente a placé le Baurubatrachus du Crétacé supérieur dans les deux genres calyptocephalellidés récents Calyptocephalella et Telmatobufo . Bien que Muzzopappa et Báez mentionnent que Calyptocephalella et Telmatobufo sont caractérisés par un neurocrâne fortement ossifié, nous ne pouvons le confirmer que pour le premier genre (Fig. 4a, f ). Chez Calyptocephalella, le motif d’ornementation des os du crâne est variable. Chez C. conquella , il est construit soit par réseau de ponctuations chez les petits individus, soit par ornementation tuberculée chez les adultes. Chez C. satan et C. casamayorensis , les os du crâne ornés sont légèrement plus grands que NRM-PZ B281 mais ils ont un motif similaire construit de fosses. C. pichifleufensis est connu par des individus plus gros qui présentent des motifs d’ornementation similaires mais avec des fosses plus grandes. Par rapport à ces espèces, la grenouille antarctique présente une ornementation très similaire à celle de C. satan et C. casamayorensis . Compte tenu de notre comparaison, nous concluons que le fragment d’os orné NRM-PZ B281 représente un os du crâne (très probablement un nasal) d’un Calyptocephalella ou Baurubatrachus de petite taille . Etant donné la présence d’une petite Calyptocephalella comme l’indique l’ilium dans le même affleurement de mesure de seulement quelques m 2 , il est fort probable que le spécimen NRM-PZ B281 appartienne au même genre. Un enregistrement comparable d’un ilium et d’os ornés se référant au genre Calyptocephalella a été mentionné dans Báez .
Une analyse phylogénétique récente a placé le Baurubatrachus du Crétacé supérieur dans les deux genres calyptocephalellidés récents Calyptocephalella et Telmatobufo . Bien que Muzzopappa et Báez mentionnent que Calyptocephalella et Telmatobufo sont caractérisés par un neurocrâne fortement ossifié, nous ne pouvons le confirmer que pour le premier genre (Fig. 4a, f ). Chez Calyptocephalella, le motif d’ornementation des os du crâne est variable. Chez C. conquella , il est construit soit par réseau de ponctuations chez les petits individus, soit par ornementation tuberculée chez les adultes. Chez C. satan et C. casamayorensis , les os du crâne ornés sont légèrement plus grands que NRM-PZ B281 mais ils ont un motif similaire construit de fosses. C. pichifleufensis est connu par des individus plus gros qui présentent des motifs d’ornementation similaires mais avec des fosses plus grandes. Par rapport à ces espèces, la grenouille antarctique présente une ornementation très similaire à celle de C. satan et C. casamayorensis . Compte tenu de notre comparaison, nous concluons que le fragment d’os orné NRM-PZ B281 représente un os du crâne (très probablement un nasal) d’un Calyptocephalella ou Baurubatrachus de petite taille . Etant donné la présence d’une petite Calyptocephalella comme l’indique l’ilium dans le même affleurement de mesure de seulement quelques m 2 , il est fort probable que le spécimen NRM-PZ B281 appartienne au même genre. Un enregistrement comparable d’un ilium et d’os ornés se référant au genre Calyptocephalella a été mentionné dans Báez .
Discussion : Parmi les amphibiens récents, les grenouilles (Anura) ont la distribution la plus large, couvrant tous les continents à l’exception de l’Antarctique, où les conditions sont inhabitables depuis plus de dizaines de millions d’années. Contrairement à tous les autres continents, aucune trace d’aucun groupe d’amphibiens existant, appartenant tous au clade des lissamphibiens, n’a été documentée en Antarctique. Cet article présente le premier enregistrement d’un lissamphibien en Antarctique, avec des fossiles de l’Éocène faisant référence à l’ordre Anura, et très probablement au genre australobatrachien Calyptocephalella .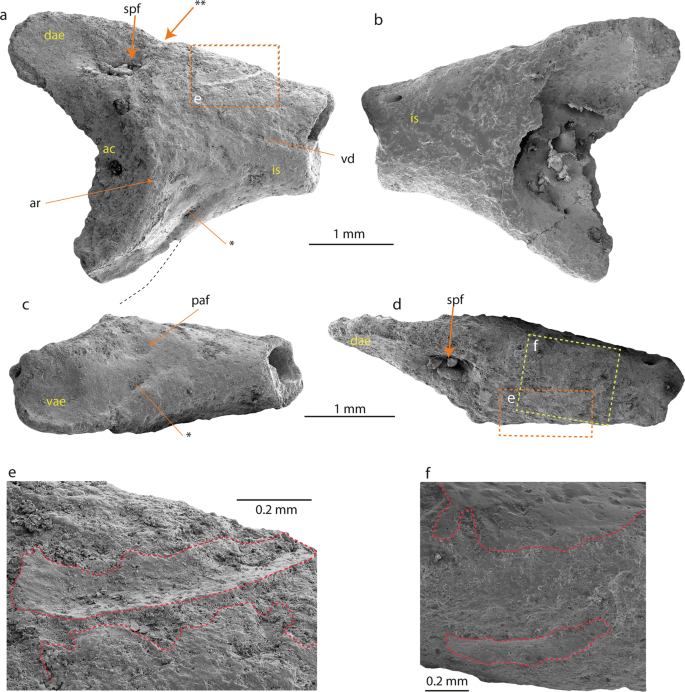 La famille des Calyptocephalellidae appartient aux grenouilles néobatraciennes et est exclusivement connue d’Amérique du Sud . Les cinq espèces existantes, y compris le genre monospécifique Calyptocephalellaavec des os crâniens hyperossifiés, sont limités aux Andes chiliennes tandis que la plupart des représentants fossiles sont connus de la Patagonie argentine. Aujourd’hui, Calyptocephalella habite les basses terres du centre du Chili (limite d’altitude supérieure à 500 m) à l’est des Andes dans des climats tempérés et humides, entre les latitudes 30 et 43 ° S. Il a un mode de vie aquatique ou semi-aquatique et peuple les masses d’eau stagnantes ou à débit lent (lacs, étangs, ruisseaux) dans les forêts tempérées de Nothofagus de Valdivian .
La famille des Calyptocephalellidae appartient aux grenouilles néobatraciennes et est exclusivement connue d’Amérique du Sud . Les cinq espèces existantes, y compris le genre monospécifique Calyptocephalellaavec des os crâniens hyperossifiés, sont limités aux Andes chiliennes tandis que la plupart des représentants fossiles sont connus de la Patagonie argentine. Aujourd’hui, Calyptocephalella habite les basses terres du centre du Chili (limite d’altitude supérieure à 500 m) à l’est des Andes dans des climats tempérés et humides, entre les latitudes 30 et 43 ° S. Il a un mode de vie aquatique ou semi-aquatique et peuple les masses d’eau stagnantes ou à débit lent (lacs, étangs, ruisseaux) dans les forêts tempérées de Nothofagus de Valdivian .
Les fossiles les plus anciens faisant référence à Calyptocephalella sont connus du Crétacé supérieur d’Argentine . Au début du Paléocène-terminal du Miocène, leur aire de répartition géographique était limitée à la Patagonie à l’est des Andes . Ce n’est qu’à la fin du Pléistocène qu’ils sont apparus à l’ouest des Andes, où ils ont leur distribution endémique actuelle .
Le clade Australobatrachia comprend les Myobatrachoidea (familles Myobatrachidae + Limnodynastidae sensu ), aujourd’hui répartis en Australie et au sud de la Nouvelle-Guinée, et la famille des Calyptocephalellidae (Batrachophrynidae sensu ). Les australobatrachies sont considérées comme un groupe souche des Hyloidea. Le myobatrachoïde le plus ancien d’Australie est au moins aussi vieux que le début de l’Éocène, sur la base d’ilia fragmentaires qui ont été référés au Lechriodus existant à la base . La scission entre les Calyptocephalellidae et les Myobatrachidae (Calyptocephalellidae + Myobatrachoidea sensu ) s’est produite à environ 100 Ma (~ limite du Crétacé précoce-tardif) .  Considérant les distributions d’Australobatrachie existantes (Fig. 5), les premiers enregistrements fossiles et l’âge de divergence (d’après les données génétiques) des lignées Calyptocephalellidae et Myobatrachoidea, il est clair que l’Antarctique a joué un rôle paléobiogéographique important pour les Australobatrachia et leur dispersion conséquente. Étant donné que l’ancêtre commun le plus récent du clade, y compris Hyloidae et Myobatrachidae + Calyptocephalellidae, s’est produit en Amérique du Sud, leur origine en Amérique du Sud et leur dispersion consécutive de l’Amérique du Sud à l’Australie via l’Antarctique ont été suggérées . De plus, cela suggère un autre cas de fortes affinités faunistiques du continent avec l’Amérique du Sud et l’Australie . Jusqu’à présent, l’Antarctique a été considérée comme une voie de dispersion, mais pas comme un lieu d’origine probable. Les nouvelles découvertes de fossiles soutiennent l’hypothèse selon laquelle l’Antarctique pourrait avoir agi comme un centre de diversification pour les australobatrachiens.
Considérant les distributions d’Australobatrachie existantes (Fig. 5), les premiers enregistrements fossiles et l’âge de divergence (d’après les données génétiques) des lignées Calyptocephalellidae et Myobatrachoidea, il est clair que l’Antarctique a joué un rôle paléobiogéographique important pour les Australobatrachia et leur dispersion conséquente. Étant donné que l’ancêtre commun le plus récent du clade, y compris Hyloidae et Myobatrachidae + Calyptocephalellidae, s’est produit en Amérique du Sud, leur origine en Amérique du Sud et leur dispersion consécutive de l’Amérique du Sud à l’Australie via l’Antarctique ont été suggérées . De plus, cela suggère un autre cas de fortes affinités faunistiques du continent avec l’Amérique du Sud et l’Australie . Jusqu’à présent, l’Antarctique a été considérée comme une voie de dispersion, mais pas comme un lieu d’origine probable. Les nouvelles découvertes de fossiles soutiennent l’hypothèse selon laquelle l’Antarctique pourrait avoir agi comme un centre de diversification pour les australobatrachiens.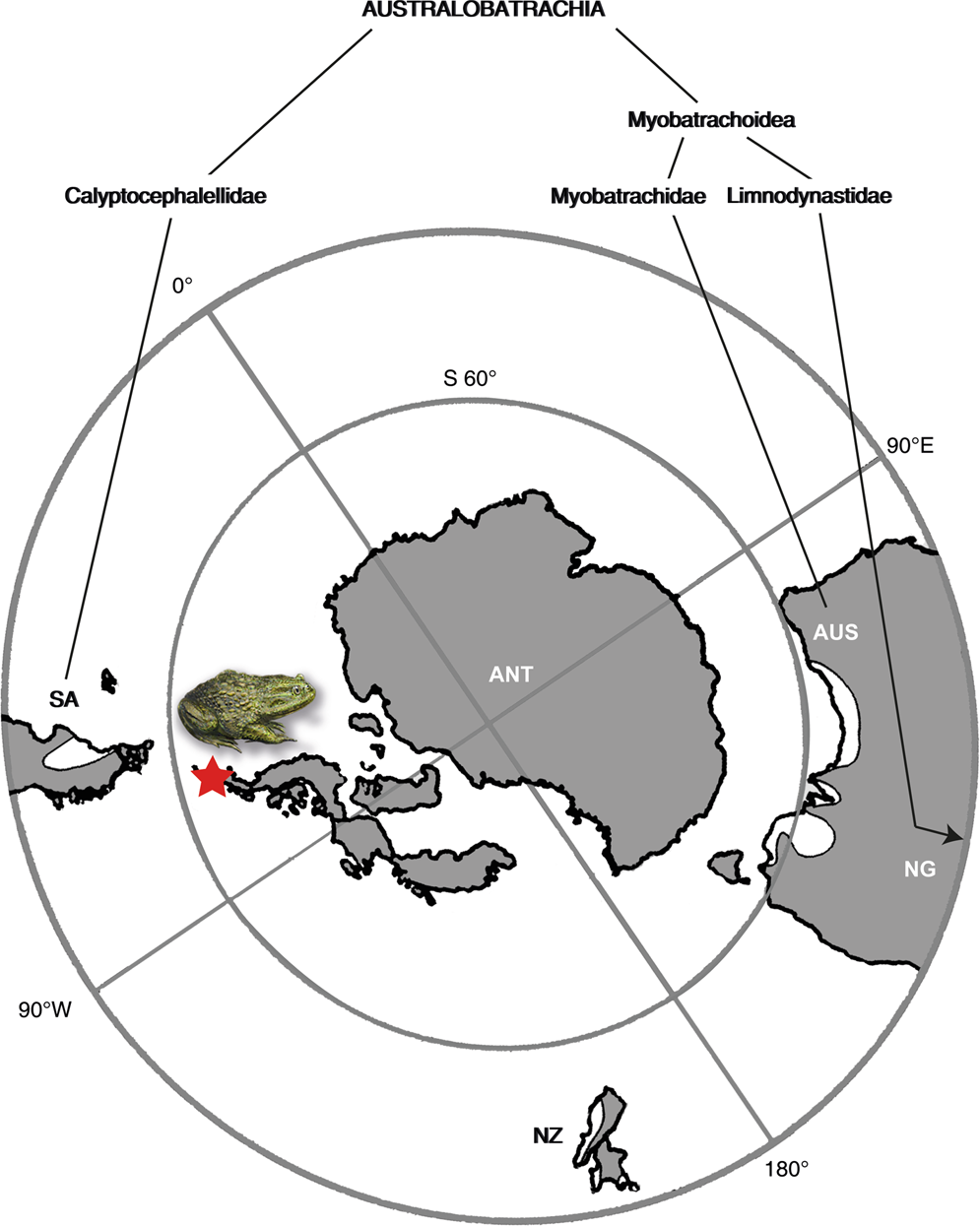 Paléogéographie éocène de la région polaire sud avec un cladogramme de grenouilles australobatrachidés montrant leurs occurrences sur les continents du sud. La couleur grise indique les contours des continents à l’Éocène, la couleur noire les contours actuels des continents. Carte redessinée à partir d’un original généré à l’aide du logiciel ArcGIS 10.17.1 ( www.esri.com ), basé sur la couche de carte de base Satellite dans google Maps (Map data ©2019 Google). Abréviations : ANT, Antarctique ; AUS, Australie et Tasmanie ; NG, Nouvelle-Guinée ; Nouvelle-Zélande, Nouvelle-Zélande ; SA, Amérique du Sud. L’étoile rouge indique la localité fossile sur l’île Seymour.
Paléogéographie éocène de la région polaire sud avec un cladogramme de grenouilles australobatrachidés montrant leurs occurrences sur les continents du sud. La couleur grise indique les contours des continents à l’Éocène, la couleur noire les contours actuels des continents. Carte redessinée à partir d’un original généré à l’aide du logiciel ArcGIS 10.17.1 ( www.esri.com ), basé sur la couche de carte de base Satellite dans google Maps (Map data ©2019 Google). Abréviations : ANT, Antarctique ; AUS, Australie et Tasmanie ; NG, Nouvelle-Guinée ; Nouvelle-Zélande, Nouvelle-Zélande ; SA, Amérique du Sud. L’étoile rouge indique la localité fossile sur l’île Seymour. Reconstitution d’une mare éocène dans la forêt de Nothofagus de la péninsule antarctique avec Calyptocephalella , reposant sur une feuille de Notonuphar antarctica qui a été décrite de la même localité . Oeuvre de Pollyanna von Knorring, Musée suédois d’histoire naturelle. Crédits photo : Simon Pierre Barrette et José Grau de Puerto Montt, Wikimedia Commons (CC BY-SA 3.0), et Mats Wedin, Musée suédois d’histoire naturelle.
Reconstitution d’une mare éocène dans la forêt de Nothofagus de la péninsule antarctique avec Calyptocephalella , reposant sur une feuille de Notonuphar antarctica qui a été décrite de la même localité . Oeuvre de Pollyanna von Knorring, Musée suédois d’histoire naturelle. Crédits photo : Simon Pierre Barrette et José Grau de Puerto Montt, Wikimedia Commons (CC BY-SA 3.0), et Mats Wedin, Musée suédois d’histoire naturelle.

La grenouille de l’île Seymour signalée ici est le premier vertébré indicatif d’habitats d’eau douce sur la péninsule antarctique de l’Éocène, à la suite de preuves d’invertébrés et de plantes (Fig. 6 ). Il est intéressant de noter que presque toutes les localités fossiles où se trouve Calyptocephalella (à l’exception de celles pour lesquelles les données sur les plantes fossiles ne sont pas disponibles) contiennent des preuves de la présence de Nothofagus , y compris l’île Seymour . Le sud de l’aire de répartition existante de Calyptocephalella se produit en sympatrie avec le marsupial microbiotherian Dromiciops gliroides (Fig. S4), également connu sous le nom de «Monito del Monte» ou «Colocolo Opossum», un petit mammifère au mode de vie arboricole et à la distribution endémique dans les forêts denses de Nothofagus valdiviens des hautes terres d’Argentine et du Chili .  Le climat de cette zone forestière de Nothofagus avec les occurrences sympatriques de ces deux animaux endémiques montre des conditions humides et tempérées (pour les valeurs numériques, voir Méthodes et Tableau 2 , Fig. S4 ). Dromiciops gliroides est la seule espèce existante de l’ordre Microbiotheria et est considérée comme le seul représentant sud-américain du super-ordre Australidelphia qui comprend autrement les marsupiaux australiens. À partir de la même petite lentille riche en coquillages qui a produit les restes de grenouilles rapportés ici, le microbiotherian fossile Woodburnodon casei a été décrit. Par conséquent, nous émettons l’hypothèse que les conditions climatiques de la péninsule antarctique au cours du Bartonien (fin de l’Éocène moyen) devraient être comparables au climat que l’on trouve aujourd’hui dans l’aire de répartition simultanée des forêts de Nothofagus habitées par Calyptocephalella et Dromiciops d’ Amérique du Sud .
Le climat de cette zone forestière de Nothofagus avec les occurrences sympatriques de ces deux animaux endémiques montre des conditions humides et tempérées (pour les valeurs numériques, voir Méthodes et Tableau 2 , Fig. S4 ). Dromiciops gliroides est la seule espèce existante de l’ordre Microbiotheria et est considérée comme le seul représentant sud-américain du super-ordre Australidelphia qui comprend autrement les marsupiaux australiens. À partir de la même petite lentille riche en coquillages qui a produit les restes de grenouilles rapportés ici, le microbiotherian fossile Woodburnodon casei a été décrit. Par conséquent, nous émettons l’hypothèse que les conditions climatiques de la péninsule antarctique au cours du Bartonien (fin de l’Éocène moyen) devraient être comparables au climat que l’on trouve aujourd’hui dans l’aire de répartition simultanée des forêts de Nothofagus habitées par Calyptocephalella et Dromiciops d’ Amérique du Sud .
Les découvertes fossiles d’une grenouille et d’un marsupial de l’île Seymour, ainsi que leurs distributions fossiles et récentes, représentent des exemples exceptionnels du rôle du changement climatique mondial sur le changement des aires biogéographiques. Malgré le refroidissement global et la disparition des habitats de ces groupes sur de vastes zones allant de l’Antarctique à la Patagonie, ils ont maintenu leur présence relictuelle dans les forêts de Nothofagus des Andes centrales chiliennes. Ainsi, la forêt valdivienne Nothofagus est un environnement unique offrant des habitats non seulement pour les réfugiés antarctiques de l’Éocène, mais fournit également un analogue moderne du climat antarctique juste avant la glaciation du continent sud.
Méthodes – Documentation : Le matériau fossile étudié a été monté sur des talons en aluminium, recouvert d’or et imagé à l’aide d’un microscope électronique à balayage à émission de champ Hitachi S-4300 au Musée suédois d’histoire naturelle (Stockholm). Pour la comparaison avec les familles concernées (tableau S1 ), les données CT du site Morphosource ont été utilisées. La visualisation et la segmentation du matériel osseux ont été réalisées à l’aide du logiciel Amira 9.0 à Porrentruy, Suisse. Nomenclature : Sauf indication contraire, la nomenclature ostéologique de cette étude suit celle de Gómez et Turazzini 66 pour la description des restes fossiles.
Nomenclature : Sauf indication contraire, la nomenclature ostéologique de cette étude suit celle de Gómez et Turazzini 66 pour la description des restes fossiles.
Estimation de la taille du corps : Les valeurs de la longueur museau-évent (SVL) de Calyptocephalella sp. de l’Antarctique ont été reconstitués à partir de photographies du squelette de C. pichileufensis et du modèle 3D de C. gayi (Tableau 1 , Fig. S3 ). La hauteur de la transition (HT) entre la diaphyse iliaque et le corps iliaque (Fig. S3 ) a été utilisée comme référence de comparaison pour reconstruire la taille corporelle du NRM-PZ B282. Le rapport du HT au SVL a été utilisé comme référence pour calculer la valeur de la longueur du museau-évent du NRM-PZ B282 individuel.
Analyse climatique : Nous avons analysé les paramètres climatiques de stations sélectionnées de la zone avec présence sympatrique de Dromiciops gliroides et Calyptocephalella gayi (Tableau 2 et Fig. S4 ). Étant donné que la limite d’altitude supérieure pour la répartition actuelle de Calyptocephalella est de 500 m au-dessus du niveau de la mer, seules les stations jusqu’à cette altitude ont été prises en compte pour l’analyse climatique. Cette analyse montre un espace climatique remarquable avec des précipitations annuelles moyennes ≥ 900 mm, une température moyenne du mois le plus froid ≥ 3,75 °C et une température moyenne du mois le plus chaud ≥ 13,79 °C (Tableau 2). Les variations des paramètres climatiques résultent à la fois des différences d’altitude et de latitude, de sorte que la température augmente et les précipitations diminuent vers le nord, alors qu’une tendance inverse est observable à des altitudes plus élevées. Les tétrapodes continentaux ectothermiques cénozoïques (amphibiens et reptiles) n’ont pas été documentés auparavant de l’Antarctique, contrairement à tous les autres continents. Nous rapportons ici un ilium fossile et un os de crâne orné qui peuvent être attribués à la famille récente, sud-américaine, des anoures Calyptocephalellidae ou grenouilles casquées, représentant le premier amphibien moderne trouvé en Antarctique. Les deux fragments d’os ont été récupérés dans des sédiments de l’Éocène, vieux d’environ 40 millions d’années, sur l’île Seymour, dans la péninsule antarctique. L’enregistrement de grenouilles calyptocéphales hyperossifiées en dehors de l’Amérique du Sud soutient le cosmopolitisme gondwanien du clade anoure Australobatrachia. Nos résultats démontrent que les écosystèmes d’eau douce de l’Éocène en Antarctique ont fourni des habitats favorables aux vertébrés ectothermes (avec des précipitations annuelles moyennes ≥ 900 mm, une température moyenne du mois le plus froid ≥ 3,75 ° C et une température moyenne du mois le plus chaud ≥ 13, 79 ° C), à un moment où il y avait au moins des calottes glaciaires éphémères existant sur les hautes terres à l’intérieur du continent.
Les tétrapodes continentaux ectothermiques cénozoïques (amphibiens et reptiles) n’ont pas été documentés auparavant de l’Antarctique, contrairement à tous les autres continents. Nous rapportons ici un ilium fossile et un os de crâne orné qui peuvent être attribués à la famille récente, sud-américaine, des anoures Calyptocephalellidae ou grenouilles casquées, représentant le premier amphibien moderne trouvé en Antarctique. Les deux fragments d’os ont été récupérés dans des sédiments de l’Éocène, vieux d’environ 40 millions d’années, sur l’île Seymour, dans la péninsule antarctique. L’enregistrement de grenouilles calyptocéphales hyperossifiées en dehors de l’Amérique du Sud soutient le cosmopolitisme gondwanien du clade anoure Australobatrachia. Nos résultats démontrent que les écosystèmes d’eau douce de l’Éocène en Antarctique ont fourni des habitats favorables aux vertébrés ectothermes (avec des précipitations annuelles moyennes ≥ 900 mm, une température moyenne du mois le plus froid ≥ 3,75 ° C et une température moyenne du mois le plus chaud ≥ 13, 79 ° C), à un moment où il y avait au moins des calottes glaciaires éphémères existant sur les hautes terres à l’intérieur du continent. Le premier fossile de grenouille de l’Antarctique a été découvert
Le premier fossile de grenouille de l’Antarctique a été découvert
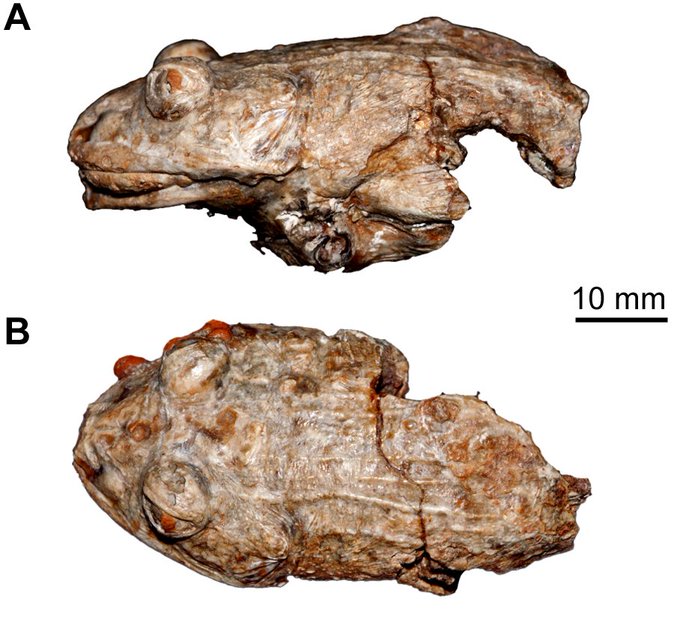
Le premier fossile de grenouille trouvé en Antarctique donne un nouvel aperçu de l’ancien climat du continent.
Les paléontologues ont découvert des fragments de l’os de la hanche et du crâne de la grenouille dans des sédiments vieux de 40 millions d’années prélevés sur l’île Seymour, près de la pointe de la péninsule antarctique.
Les scientifiques ont déjà trouvé des preuves d’ amphibiens géants qui ont parcouru l’Antarctique pendant la période du Trias, il y a plus de 200 millions d’années, mais aucune trace sur le continent d’amphibiens comme ceux qui existent aujourd’hui ( SN : 23/03/15 ). La forme des ossements nouvellement découverts indique que cette grenouille appartenait à la famille des Calyptocephalellidae , ou grenouilles casquées, que l’on trouve aujourd’hui en Amérique du Sud. Les parents modernes de la grenouille fossilisée vivent exclusivement dans les Andes centrales chiliennes chaudes et humides. Cela suggère que des conditions climatiques similaires existaient en Antarctique il y a environ 40 millions d’années, rapportent des chercheurs le 23 avril dans Scientific Reports .
Les parents modernes de la grenouille fossilisée vivent exclusivement dans les Andes centrales chiliennes chaudes et humides. Cela suggère que des conditions climatiques similaires existaient en Antarctique il y a environ 40 millions d’années, rapportent des chercheurs le 23 avril dans Scientific Reports .
Cela donne un indice sur la rapidité avec laquelle l’Antarctique est passé d’un froid doux à un froid glacial ( SN : 4/1/20 ). L’Antarctique a rapidement gelé après s’être séparé de l’Australie et de l’Amérique du Sud, qui faisaient autrefois partie du supercontinent Gondwana ( SN : 10/10/19 ). Mais certaines preuves géologiques suggèrent que les calottes glaciaires ont commencé à se former sur l’Antarctique avant qu’il ne se sépare complètement des autres continents du sud il y a environ 34 millions d’années. « La question est maintenant, à quel point faisait-il froid et qu’est-ce qui vivait sur le continent lorsque ces calottes glaciaires ont commencé à se former? » explique le co-auteur de l’étude, Thomas Mörs, paléontologue au Musée suédois d’histoire naturelle de Stockholm. « Cette grenouille est une indication de plus qu’à [cette] époque, du moins autour de la péninsule, c’était encore un habitat convenable pour les animaux à sang froid comme les reptiles et les amphibiens. »
« La question est maintenant, à quel point faisait-il froid et qu’est-ce qui vivait sur le continent lorsque ces calottes glaciaires ont commencé à se former? » explique le co-auteur de l’étude, Thomas Mörs, paléontologue au Musée suédois d’histoire naturelle de Stockholm. « Cette grenouille est une indication de plus qu’à [cette] époque, du moins autour de la péninsule, c’était encore un habitat convenable pour les animaux à sang froid comme les reptiles et les amphibiens. »
https://www.courthousenews.com/frog-fossils-in-antarctica-offer-evidence-of-warmer-past/
https://www.nature.com/articles/s41598-020-61973-5