 Un crâne complet de Dmanissi, en Géorgie, et la biologie évolutive des premiers Homo
Un crâne complet de Dmanissi, en Géorgie, et la biologie évolutive des premiers Homo  Quand Homo sapiens a-t-il atteint l’Asie du Sud-Est et le Sahul pour la première fois ?
Quand Homo sapiens a-t-il atteint l’Asie du Sud-Est et le Sahul pour la première fois ? Suite à une étude sur un crâne découvert en Géorgie, on remet en question la classification des premiers «Homo» en plusieurs espèces.
 Suite à une étude sur un crâne découvert en Géorgie, on remet en question la classification des premiers « Homo » en plusieurs espèces. Le crâne complet d’un premier Homo suggère une seule lignée évolutive. Le crâne d’un lointain ancêtre de l’Homme laisse penser que toutes les espèces d’Homo n’en formaient qu’une.
Suite à une étude sur un crâne découvert en Géorgie, on remet en question la classification des premiers « Homo » en plusieurs espèces. Le crâne complet d’un premier Homo suggère une seule lignée évolutive. Le crâne d’un lointain ancêtre de l’Homme laisse penser que toutes les espèces d’Homo n’en formaient qu’une.
 Une trouvaille enivrante
Une trouvaille enivrante
Au cours des deux dernières décennies, des fouilles sur le site archéologique de Dmanissi, en Géorgie, ont révélé des fossiles d’hominidés du Pléistocène le plus ancien, peu après la première dispersion du genre Homo au-delà de l’Afrique. Lordkipanidze et al. décrivent maintenant un crâne fossile du site. Combiné avec des restes mandibulaires qui avaient été trouvés plus tôt, cette découverte complète le premier crâne d’hominidé entier de cette période. Résumé
Résumé
Le site de Dmanissi, en Géorgie, a livré un échantillon impressionnant de restes crâniens et post-crâniens d’hominidés, documentant la présence d’Homo hors d’Afrique il y a environ 1,8 million d’années. Nous rapportons ici un nouveau crâne de Dmanissi (D4500) qui, avec sa mandibule (D2600), représente le premier crâne d’hominidé adulte complètement préservé au monde du début du Pléistocène. D4500/D2600 combine un petit casse-tête (546 centimètres cubes) avec un grand visage prognathique et présente des affinités morphologiques étroites avec les premiers fossiles d’Homo connus d’Afrique. L’échantillon de Dmanissi, qui comprend maintenant cinq crânes, fournit des preuves directes d’une grande variation morphologique au sein et parmi les premiers paléodèmes Homo. 
 Quand Homo sapiens a-t-il atteint l’Asie du Sud-Est et le Sahul pour la première fois ?
Quand Homo sapiens a-t-il atteint l’Asie du Sud-Est et le Sahul pour la première fois ? Les humains anatomiquement modernes (Homo sapiens, AMH) ont commencé à se répandre à travers l’Eurasie depuis l’Afrique et l’Asie du Sud-Ouest adjacente il y a environ 50 000 à 55 000 ans (environ 50 à 55 ka). Certains ont fait valoir que les données génétiques humaines, fossiles et archéologiques indiquent une ou plusieurs dispersions antérieures, peut-être dès 120 ka. Une estimation d’âge récemment rapportée de 65 ka pour Madjedbebe, un site archéologique dans le nord du Sahul (Pléistocène Australie-Nouvelle-Guinée), si elle est correcte, offre ce qui pourrait être le soutien le plus solide jamais présenté pour un exode africain d’AMH pré-55 ka.
Les humains anatomiquement modernes (Homo sapiens, AMH) ont commencé à se répandre à travers l’Eurasie depuis l’Afrique et l’Asie du Sud-Ouest adjacente il y a environ 50 000 à 55 000 ans (environ 50 à 55 ka). Certains ont fait valoir que les données génétiques humaines, fossiles et archéologiques indiquent une ou plusieurs dispersions antérieures, peut-être dès 120 ka. Une estimation d’âge récemment rapportée de 65 ka pour Madjedbebe, un site archéologique dans le nord du Sahul (Pléistocène Australie-Nouvelle-Guinée), si elle est correcte, offre ce qui pourrait être le soutien le plus solide jamais présenté pour un exode africain d’AMH pré-55 ka. Nous examinons les preuves de l’arrivée de l’AMH sur un arc s’étendant du sud de la Chine à Sahul, puis évaluons les données de Madjedbebe. Nous constatons qu’une estimation d’âge > 50 ka pour ce site est peu susceptible d’être valide. Bien que l’AMH ait pu s’être déplacé bien au-delà de l’Afrique bien avant 50-55 ka, les données de la région d’intérêt proposées à l’appui de cette idée ne sont pas convaincantes.
Nous examinons les preuves de l’arrivée de l’AMH sur un arc s’étendant du sud de la Chine à Sahul, puis évaluons les données de Madjedbebe. Nous constatons qu’une estimation d’âge > 50 ka pour ce site est peu susceptible d’être valide. Bien que l’AMH ait pu s’être déplacé bien au-delà de l’Afrique bien avant 50-55 ka, les données de la région d’intérêt proposées à l’appui de cette idée ne sont pas convaincantes. Les données fossiles suggèrent que la lignée humaine moderne est apparue en Afrique vers 300 ka. Il existe un large consensus, mais pas universel, sur le fait que les humains quasi-modernes ou modernes étaient confinés à l’Afrique et aux régions voisines de l’Asie du Sud-Ouest jusqu’à ca. 50–55 ka. Après cette période, selon l’argument, les humains anatomiquement modernes (AMH) se sont rapidement répandus à travers l’Eurasie, déplaçant et se croisant occasionnellement avec les Néandertaliens et d’autres humains archaïques et colonisant de vastes zones (Sahul et nord de la Sibérie ; plus tard les Amériques et l’Océanie) qu’aucun hominidé n’avait occupé auparavant. Ce processus est parfois appelé le modèle « à vague unique » de dispersion de l’AMH au-delà de l’Afrique.
Les données fossiles suggèrent que la lignée humaine moderne est apparue en Afrique vers 300 ka. Il existe un large consensus, mais pas universel, sur le fait que les humains quasi-modernes ou modernes étaient confinés à l’Afrique et aux régions voisines de l’Asie du Sud-Ouest jusqu’à ca. 50–55 ka. Après cette période, selon l’argument, les humains anatomiquement modernes (AMH) se sont rapidement répandus à travers l’Eurasie, déplaçant et se croisant occasionnellement avec les Néandertaliens et d’autres humains archaïques et colonisant de vastes zones (Sahul et nord de la Sibérie ; plus tard les Amériques et l’Océanie) qu’aucun hominidé n’avait occupé auparavant. Ce processus est parfois appelé le modèle « à vague unique » de dispersion de l’AMH au-delà de l’Afrique.  Au cours de la dernière décennie environ, certains analystes ont suggéré une série d’événements plus complexes commençant bien avant 50-55 ka (voir réf. 4 et 5 pour examen), un argument fondé sur des preuves archéologiques d’Asie du Sud, des restes fossiles d’Orient et d’Afrique. Asie du Sud-Est et données génétiques du sud de l’Eurasie et de l’Australie. Les défenseurs envisagent une propagation progressive en au moins deux étapes, la première dans le sud de l’Eurasie et une autre beaucoup plus tard dans des latitudes plus élevées, y compris l’Europe.
Au cours de la dernière décennie environ, certains analystes ont suggéré une série d’événements plus complexes commençant bien avant 50-55 ka (voir réf. 4 et 5 pour examen), un argument fondé sur des preuves archéologiques d’Asie du Sud, des restes fossiles d’Orient et d’Afrique. Asie du Sud-Est et données génétiques du sud de l’Eurasie et de l’Australie. Les défenseurs envisagent une propagation progressive en au moins deux étapes, la première dans le sud de l’Eurasie et une autre beaucoup plus tard dans des latitudes plus élevées, y compris l’Europe.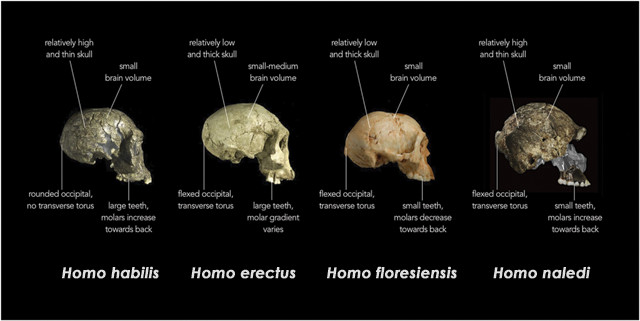 La dispersion du Pléistocène tardif depuis l’Afrique, autrefois un problème, est maintenant rarement remis en question, mais résoudre ses détails dans le temps et dans l’espace est essentiel pour l’expliquer. L’estimation d’âge de 65 ka récemment rapportée pour le site de Madjedbebe, dans le nord de l’Australie, pourrait être une contribution importante à cette discussion. Si les âges et les liens proposés avec les artefacts sont tels que rapportés, ils représenteraient l’ensemble de preuves le plus convaincant jamais proposé pour une expansion de l’AMH hors d’Afrique > 50 à 55 ka. L’âge précoce proposé pour ce site fut rapidement accepté par certains commentateurs mais contesté par d’autres. Nous examinons ici les problèmes, en particulier ceux de l’Asie du Sud-Est et des régions plus au sud et à l’est, où des données fossiles, archéologiques et génétiques critiques, ainsi que des rapports sur Madjedbebe, ont émergé au cours de la dernière décennie.
La dispersion du Pléistocène tardif depuis l’Afrique, autrefois un problème, est maintenant rarement remis en question, mais résoudre ses détails dans le temps et dans l’espace est essentiel pour l’expliquer. L’estimation d’âge de 65 ka récemment rapportée pour le site de Madjedbebe, dans le nord de l’Australie, pourrait être une contribution importante à cette discussion. Si les âges et les liens proposés avec les artefacts sont tels que rapportés, ils représenteraient l’ensemble de preuves le plus convaincant jamais proposé pour une expansion de l’AMH hors d’Afrique > 50 à 55 ka. L’âge précoce proposé pour ce site fut rapidement accepté par certains commentateurs mais contesté par d’autres. Nous examinons ici les problèmes, en particulier ceux de l’Asie du Sud-Est et des régions plus au sud et à l’est, où des données fossiles, archéologiques et génétiques critiques, ainsi que des rapports sur Madjedbebe, ont émergé au cours de la dernière décennie.  Paramètre
Paramètre
La zone d’intérêt s’étend du sud et de l’est de la Chine méridionale à travers l’Asie du Sud-Est continentale, l’archipel malais et l’Australie-Nouvelle-Guinée. Nous l’appelons ici «l’arc Chine du Sud-Sahul (SCS)». Il a eu une histoire environnementale complexe au cours des derniers millions d’années, principalement en raison des changements dans la géométrie orbitale de la Terre et de leur impact sur le climat et le niveau de la mer. Ici, nous nous concentrons sur la première partie du dernier cycle glaciaire (40-130 ka), la période de temps entourant la première preuve définitive et la plupart des premières prétendues preuves d’AMH dans la région. Au cours du dernier interglaciaire (115-130 ka), le niveau de la mer était de 6 à 10 m plus élevé qu’actuellement, mais les configurations du relief étaient globalement similaires à celles d’aujourd’hui. Par la suite, le niveau de la mer a fluctué de manière irrégulière, tombant finalement au dernier maximum glaciaire, moins 120 m, à 20 ka.
Au cours du dernier interglaciaire (115-130 ka), le niveau de la mer était de 6 à 10 m plus élevé qu’actuellement, mais les configurations du relief étaient globalement similaires à celles d’aujourd’hui. Par la suite, le niveau de la mer a fluctué de manière irrégulière, tombant finalement au dernier maximum glaciaire, moins 120 m, à 20 ka.  Chutes de 40 m ou plus exposées de grandes zones maintenant submergées des plateaux Sunda et Sahul, dans le premier cas rejoignant Sumatra, Bornéo et Java dans le cadre d’une large péninsule s’étendant au sud et à l’est de l’actuel continent sud-est asiatique et dans le second créant des liaisons terrestres sèches entre l’Australie, la Nouvelle-Guinée et la Tasmanie. Les îles de Wallacea, le vaste archipel entre Sunda et Sahul, sont restées sans lien avec l’une ou l’autre, formant la barrière entre les provinces biogéographiques orientales et australiennes reconnues à l’origine par Alfred Russel Wallace. Aucun mammifère terrestre de grande taille n’a réussi un transit complet et démographiquement réussi de ce seuil avant l’arrivée des AMH.
Chutes de 40 m ou plus exposées de grandes zones maintenant submergées des plateaux Sunda et Sahul, dans le premier cas rejoignant Sumatra, Bornéo et Java dans le cadre d’une large péninsule s’étendant au sud et à l’est de l’actuel continent sud-est asiatique et dans le second créant des liaisons terrestres sèches entre l’Australie, la Nouvelle-Guinée et la Tasmanie. Les îles de Wallacea, le vaste archipel entre Sunda et Sahul, sont restées sans lien avec l’une ou l’autre, formant la barrière entre les provinces biogéographiques orientales et australiennes reconnues à l’origine par Alfred Russel Wallace. Aucun mammifère terrestre de grande taille n’a réussi un transit complet et démographiquement réussi de ce seuil avant l’arrivée des AMH. Preuve fossile de la présence d’AMH
Preuve fossile de la présence d’AMH  Quatre espèces d’Homo sont identifiées dans la région d’intérêt. Les caractéristiques définitives comprennent des aspects de la taille du corps, de la robustesse du squelette et de l’anatomie crânienne (annexe SI, SI.1.1). Homo erectus, d’origine africaine et le plus ancien représentant définitif du genre, peut avoir été présent à Java il y a 1,5 Ma (15) et peut-être y avoir persisté et en Asie du Sud-Est plus largement jusqu’à l’arrivée de l’AMH au Pléistocène supérieur (comparer les réf. 16 et 17). Vers 1,0 Ma, H. erectus avait atteint Flores, où il a probablement donné naissance à la naine insulaire Homo floresiensis.
Quatre espèces d’Homo sont identifiées dans la région d’intérêt. Les caractéristiques définitives comprennent des aspects de la taille du corps, de la robustesse du squelette et de l’anatomie crânienne (annexe SI, SI.1.1). Homo erectus, d’origine africaine et le plus ancien représentant définitif du genre, peut avoir été présent à Java il y a 1,5 Ma (15) et peut-être y avoir persisté et en Asie du Sud-Est plus largement jusqu’à l’arrivée de l’AMH au Pléistocène supérieur (comparer les réf. 16 et 17). Vers 1,0 Ma, H. erectus avait atteint Flores, où il a probablement donné naissance à la naine insulaire Homo floresiensis.  En supposant que les configurations de relief actuelles (l’activité tectonique passée n’ayant pas été un facteur de complication), le passage sur la route la plus courte de Sunda à Flores a nécessité deux traversées océaniques ouvertes, le plus long ca. 20 km, tous deux compliqués par le fort courant traversant indonésien nord-sud. Un modèle alternatif suggère de voyager via les Philippines et Sulawasi, un itinéraire qui a nécessité plusieurs traversées (une > 80 km) mais qui peut avoir été facilité par ce même courant nord-sud. Quel que soit l’itinéraire emprunté, le transit est considéré comme accidentel. Archaïque Homo sapiens, distingué par des tailles de cerveau dans la gamme humaine moderne mais par ailleurs largement similaire à la fin de H. erectus, était présent dans le sud de la Chine et vers le sud de 100 à 400 ka ou plus tard et peut avoir inclus des représentants de la lignée définie sur des bases génomiques comme Denisovan. Les gènes de Denisovan ont été détectés à de faibles niveaux dans les populations humaines vivantes d’Asie du Sud et du Pacifique et à des niveaux relativement élevés chez les Australo-Mélanésiens.
En supposant que les configurations de relief actuelles (l’activité tectonique passée n’ayant pas été un facteur de complication), le passage sur la route la plus courte de Sunda à Flores a nécessité deux traversées océaniques ouvertes, le plus long ca. 20 km, tous deux compliqués par le fort courant traversant indonésien nord-sud. Un modèle alternatif suggère de voyager via les Philippines et Sulawasi, un itinéraire qui a nécessité plusieurs traversées (une > 80 km) mais qui peut avoir été facilité par ce même courant nord-sud. Quel que soit l’itinéraire emprunté, le transit est considéré comme accidentel. Archaïque Homo sapiens, distingué par des tailles de cerveau dans la gamme humaine moderne mais par ailleurs largement similaire à la fin de H. erectus, était présent dans le sud de la Chine et vers le sud de 100 à 400 ka ou plus tard et peut avoir inclus des représentants de la lignée définie sur des bases génomiques comme Denisovan. Les gènes de Denisovan ont été détectés à de faibles niveaux dans les populations humaines vivantes d’Asie du Sud et du Pacifique et à des niveaux relativement élevés chez les Australo-Mélanésiens. Dix-sept sites dans la zone et la période d’intérêt ont livré des restes humains classés par au moins certains analystes comme anatomiquement modernes (l’annexe SI, SI.1.2 fournit des détails). Neuf de ces sites se trouvent dans le sud de la Chine ; huit sont plus au sud et à l’est. Comme publié jusqu’à présent, 9 des 17 sites n’ont fourni que des dents, et sept de ces sites n’ont produit qu’une ou deux dents. Un site a produit un squelette complet ; cinq ont livré des crânes isolés, des mandibules ou des fragments de ceux-ci ; deux sites ont fourni des éléments post-crâniens uniques. Les âges fossiles estimés vont de 36 à 220 ka ; la plupart tombent dans l’intervalle 40–120 ka. La plupart des estimations d’âge sont basées sur des analyses de la série U de pierres d’écoulement censées encadrer les restes humains. Plusieurs ont été réalisés à partir d’analyses des restes eux-mêmes. D’autres techniques de datation employées comprennent le 14C, la résonance de spin électronique, la luminescence et les corrélations fauniques.
Dix-sept sites dans la zone et la période d’intérêt ont livré des restes humains classés par au moins certains analystes comme anatomiquement modernes (l’annexe SI, SI.1.2 fournit des détails). Neuf de ces sites se trouvent dans le sud de la Chine ; huit sont plus au sud et à l’est. Comme publié jusqu’à présent, 9 des 17 sites n’ont fourni que des dents, et sept de ces sites n’ont produit qu’une ou deux dents. Un site a produit un squelette complet ; cinq ont livré des crânes isolés, des mandibules ou des fragments de ceux-ci ; deux sites ont fourni des éléments post-crâniens uniques. Les âges fossiles estimés vont de 36 à 220 ka ; la plupart tombent dans l’intervalle 40–120 ka. La plupart des estimations d’âge sont basées sur des analyses de la série U de pierres d’écoulement censées encadrer les restes humains. Plusieurs ont été réalisés à partir d’analyses des restes eux-mêmes. D’autres techniques de datation employées comprennent le 14C, la résonance de spin électronique, la luminescence et les corrélations fauniques.
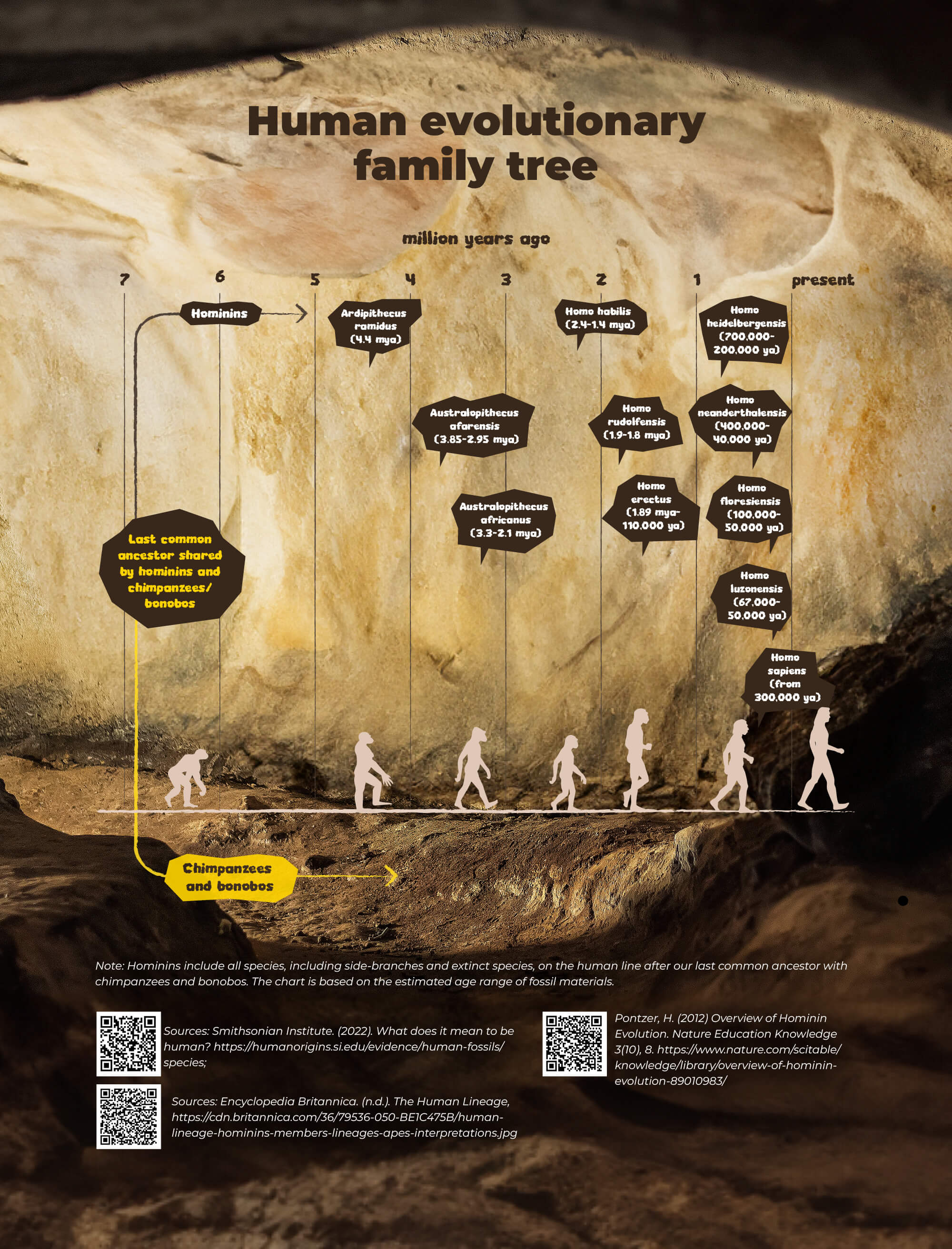
Crâne de Dmanissi : Homo Habilis, Homo Erectus appartenaient à la même espèce – Étude Le cinquième crâne découvert à Dmanissi présente une combinaison de caractéristiques inconnues des chercheurs avant la découverte – le plus grand visage, la mâchoire et les dents les plus massivement construites et le plus petit cerveau du groupe de Dmanissi. Auparavant, quatre crânes d’hominidés également bien conservés ainsi que quelques parties de squelettes y avaient été retrouvés. Dans l’ensemble, les découvertes montrent que les premiers représentants du genre Homo ont commencé à s’étendre de l’Afrique à l’Eurasie il y a 1 850 000 ans. Parce que le crâne est complètement intact, il peut fournir des réponses à diverses questions qui jusqu’à présent offraient un large champ de spéculation – à savoir, le début évolutif du genre Homo en Afrique il y a environ deux millions d’années au début de l’ère glaciaire, également appelé Pléistocène. Le débat est de savoir s’il y avait plusieurs espèces d’Homo spécialisées en Afrique à l’époque, dont au moins une était capable de se propager également en dehors de l’Afrique, ou une seule espèce capable de faire face à une variété d’écosystèmes.
Le cinquième crâne découvert à Dmanissi présente une combinaison de caractéristiques inconnues des chercheurs avant la découverte – le plus grand visage, la mâchoire et les dents les plus massivement construites et le plus petit cerveau du groupe de Dmanissi. Auparavant, quatre crânes d’hominidés également bien conservés ainsi que quelques parties de squelettes y avaient été retrouvés. Dans l’ensemble, les découvertes montrent que les premiers représentants du genre Homo ont commencé à s’étendre de l’Afrique à l’Eurasie il y a 1 850 000 ans. Parce que le crâne est complètement intact, il peut fournir des réponses à diverses questions qui jusqu’à présent offraient un large champ de spéculation – à savoir, le début évolutif du genre Homo en Afrique il y a environ deux millions d’années au début de l’ère glaciaire, également appelé Pléistocène. Le débat est de savoir s’il y avait plusieurs espèces d’Homo spécialisées en Afrique à l’époque, dont au moins une était capable de se propager également en dehors de l’Afrique, ou une seule espèce capable de faire face à une variété d’écosystèmes.
Les premières découvertes d’Homo en Afrique ont démontré une grande variation, mais il n’a pas été possible de se prononcer sur les réponses à ces questions. Christoph Zollikofer, anthropologue à l’Université de Zurich, explique pourquoi les preuves fossiles n’étaient pas suffisantes. « La plupart de ces fossiles représentent des découvertes fragmentaires uniques provenant de plusieurs points dans l’espace et dans le temps géologique d’au moins 500 000 ans. Cela rend finalement difficile la reconnaissance des variations entre les espèces dans les fossiles africains par opposition aux variations au sein des espèces. » Ce que le nouveau fossile signifie, disent-ils, c’est que les premiers membres de notre genre Homo – Homo habilis, Homo rudolfensis, Homo erectus et ainsi de suite – appartenaient en fait à la même espèce et avaient simplement l’air différent les uns des autres.
Autant d’espèces que de chercheurs 
Marcia Ponce de León, également anthropologue à l’Université de Zurich, indique une autre raison : les paléoanthropologues supposaient souvent tacitement que le fossile qu’ils venaient de trouver était représentatif de l’espèce, c’est-à-dire qu’il démontrait bien les caractéristiques de l’espèce. Statistiquement, ce n’est pas très probable, dit-elle, mais des chercheurs ont néanmoins proposé jusqu’à cinq espèces contemporaines d’Homo précoces en Afrique, notamment Homo habilis, Homo rudolfensis, Homo ergaster et Homo erectus. Ponce de León résume le problème comme suit : « Actuellement, il y a autant de subdivisions entre les espèces qu’il y a de chercheurs examinant ce problème. Dmanissi offre la clé de la solution, disent les auteurs. Selon Zollikofer, la raison pour laquelle Skull 5 est si important est qu’il réunit des caractéristiques qui ont été utilisées auparavant comme argument pour définir différentes espèces africaines.  En d’autres termes, « si le casse-tête et le visage de l’échantillon de Dmanissi avaient été trouvés comme des fossiles distincts, ils auraient très probablement été attribués à deux espèces différentes. » Ponce de León ajoute : « Il est également décisif que nous ayons cinq individus bien conservés à Dmanissi dont nous savons qu’ils ont vécu au même endroit et au même moment ».
En d’autres termes, « si le casse-tête et le visage de l’échantillon de Dmanissi avaient été trouvés comme des fossiles distincts, ils auraient très probablement été attribués à deux espèces différentes. » Ponce de León ajoute : « Il est également décisif que nous ayons cinq individus bien conservés à Dmanissi dont nous savons qu’ils ont vécu au même endroit et au même moment ».  Ces circonstances uniques de la découverte permettent de comparer la variation à Dmanissi avec la variation des populations humaines et de chimpanzés modernes. Zollikofer résume le résultat des analyses statistiques : « Premièrement, les individus de Dmanissi appartiennent tous à une population d’une seule espèce d’Homo précoce. Deuxièmement, les cinq individus de Dmanissi sont manifestement différents les uns des autres, mais pas plus différents que cinq individus humains modernes, ou cinq individus de chimpanzés d’une population donnée. »
Ces circonstances uniques de la découverte permettent de comparer la variation à Dmanissi avec la variation des populations humaines et de chimpanzés modernes. Zollikofer résume le résultat des analyses statistiques : « Premièrement, les individus de Dmanissi appartiennent tous à une population d’une seule espèce d’Homo précoce. Deuxièmement, les cinq individus de Dmanissi sont manifestement différents les uns des autres, mais pas plus différents que cinq individus humains modernes, ou cinq individus de chimpanzés d’une population donnée. »
La diversité au sein d’une espèce est donc la règle plutôt que l’exception. Les présents résultats sont étayés par une étude supplémentaire (voir la deuxième citation ci-dessous). Dans cette étude, Ponce de León, Zollikofer et d’autres collègues montrent que les différences de morphologie de la mâchoire entre les individus de Dmanissi sont principalement dues à des différences d’usure dentaire. Cela montre la nécessité d’un changement de perspective : les fossiles africains datant d’environ 1,8 million d’années représentent probablement des représentants d’une seule et même espèce, mieux décrite comme Homo erectus.  Cela suggérerait que l’Homo erectus a évolué il y a environ 2 millions d’années en Afrique, et s’est rapidement étendu à travers l’Eurasie – via des endroits tels que Dmanissi – jusqu’à la Chine et Java, où il est documenté pour la première fois il y a environ 1,2 million d’années. La comparaison des modèles de diversité en Afrique, en Eurasie et en Asie de l’Est fournit des indices sur la biologie des populations de cette première espèce humaine mondiale.
Cela suggérerait que l’Homo erectus a évolué il y a environ 2 millions d’années en Afrique, et s’est rapidement étendu à travers l’Eurasie – via des endroits tels que Dmanissi – jusqu’à la Chine et Java, où il est documenté pour la première fois il y a environ 1,2 million d’années. La comparaison des modèles de diversité en Afrique, en Eurasie et en Asie de l’Est fournit des indices sur la biologie des populations de cette première espèce humaine mondiale.
Cela fait d’Homo erectus le premier acteur mondial de l’évolution humaine. Sa redéfinition offrirait l’opportunité de suivre cette espèce humaine fossile sur une durée de 1 million d’années.
Homo sapiens et ses contemporains : les populations humaines du Pléistocène moyen et supérieur Dernière représentante de la sous-tribu des hominines, notre espèce, Homo sapiens, est l’objet de nombreux débats touchant à son origine, sa définition, sa diversité et ses relations avec les autres taxons d’hominines plus anciens ou contemporains. À travers la revue des données paléoanthropologiques et génétiques dont nous disposons actuellement, il est possible de discuter du degré de parenté et d’interaction durant le Pléistocène moyen et supérieur entre les différents représentants de notre espèce et leurs contemporains eurasiatiques dont font partie, entre autres, les Néandertaliens et les Dénisoviens.
Dernière représentante de la sous-tribu des hominines, notre espèce, Homo sapiens, est l’objet de nombreux débats touchant à son origine, sa définition, sa diversité et ses relations avec les autres taxons d’hominines plus anciens ou contemporains. À travers la revue des données paléoanthropologiques et génétiques dont nous disposons actuellement, il est possible de discuter du degré de parenté et d’interaction durant le Pléistocène moyen et supérieur entre les différents représentants de notre espèce et leurs contemporains eurasiatiques dont font partie, entre autres, les Néandertaliens et les Dénisoviens.
https://www.pnas.org/doi/10.1073/pnas.1808385115