


🧠 FUN FACT🧠
The 1963 Nobel Prize for Physiology or Medicine was awarded to Sir John Eccles, Alan Lloyd Hodgkin & Andrew Fielding Huxley for their work on ionic mechanisms involved in excitation & inhibition in the peripheral & central portion of the nerve cell membrane.🏆🤓 pic.twitter.com/QsSyVkdHww
— ISNR (@ISNRORG) January 22, 2024
Andrew a fait ses études à la University College School (1925-1930) et à la Westminster School (1930-1935, King’s Scholar); et monta au Trinity College de Cambridge en 1935 avec une bourse d’entrée majeure. Il s’était tourné vers la science à partir des classiques en 1932 et était allé à Cambridge en espérant que sa carrière serait dans les sciences physiques : il a toujours eu l’esprit mécanique et il s’est inspiré à Westminster de l’enseignement de la physique de feu JF Rudwick. Il a naturellement suivi la physique, la chimie et les mathématiques dans sa partie I à Cambridge, mais les règles l’obligeaient à choisir une autre science et il a choisi la physiologie, en grande partie sur la recommandation d’un vieil ami, B. Delisle Burns, maintenant du département de physiologie de McGill. Université. Huxley a trouvé la physiologie intéressante, en partie pour son sujet et en partie par le contact avec Adrian , Roughton, Rushton, Hodgkin et feu GA Millikan (tous boursiers de Trinity) et d’autres dans le département, et il a décidé de se spécialiser dans ce domaine. Il a passé 1937-1938 à faire de l’anatomie avec l’intention de se qualifier en médecine, et 1938-1939 à suivre le cours de la partie II en physiologie. En août 1939, Huxley rejoignit Hodgkin au Laboratoire de biologie marine de Plymouth pour sa première introduction à la recherche, et ils réussirent à enregistrer électriquement depuis l’intérieur de l’axone géant du calmar.
Huxley a été rédacteur en chef du Journal of Physiology de 1950 à 1957, il a également été rédacteur en chef du Journal of Molecular Biology. Il est devenu membre de la Royal Society en 1955 et a siégé à son conseil de 1960 à 1962. Il a travaillé à Woods Hole, Massachusetts, en 1953 en tant que boursier Lalor ; a donné les conférences Herter à la Johns Hopkins Medical School en 1959; et les conférences Jesup à l’Université de Columbia en 1964.
En 1947, Andrew Huxley épousa Jocelyn Richenda Gammell Pease, fille de MS Pease, généticienne, et de l’hon. HB Pease (née Wedgwood). Mme Huxley est juge de paix et est active dans divers travaux publics dans le Cambridgeshire. Ils ont six enfants: Janet Rachel (née le 20 avril 1948), qui a étudié la biochimie à l’Université de Bristol ; Stewart Leonard (né le 19 décembre 1949), qui a étudié l’ingénierie à l’Université de Cambridge ; Camilla Rosalind (née le 12 mars 1952), entrée à l’Université de Cambridge en 1971; Eleanor Bruce (née le 21 février 1959); Henrietta Catherine (née le 25 décembre 1960); et Clare Marjory Pease (née le 4 novembre 1962).
Affiliation au moment de l’attribution : University College, Londres, Royaume-Uni
Ses travaux : Le système nerveux des humains et des animaux est constitué de nombreuses cellules différentes. Dans les cellules, les signaux sont véhiculés par de petits courants électriques et par des substances chimiques. En mesurant les changements de charges électriques dans une très grande fibre nerveuse d’une espèce de poulpe, Andrew Huxley et Alan Hodgkin ont pu montrer comment les impulsions nerveuses sont échangées entre les cellules. En 1952, ils ont pu démontrer qu’un mécanisme fondamental implique le passage d’ions sodium et potassium dans des directions opposées à l’intérieur et à l’extérieur de la paroi cellulaire, ce qui donne lieu à des charges électriques.
Andrew Fielding Huxley (1917-2012) Andrew Fielding Huxley, OM 1983; Kt 1974; FRS 1955; PRS 1980–1985, MA, Hon. ScD Cantab. Physiologiste et biophysicien. Né à Hampstead, Londres, le 22 novembre 1917. Démonstrateur, 1946–50, directeur adjoint de la recherche, 1951–59, et lecteur en biophysique expérimentale, 1959–60, Physiological Laboratory, Cambridge ; directeur des études, Trinity College, Cambridge, 1952-1960 ; Professeur Jodrell 1960–69, professeur de recherche de la Royal Society, 1969–83, University College London. Master, 1984–90, Fellow, 1941–60 et depuis 1990, Trinity College, Cambridge (Hon. Fellow, 1967–90). Décédé le 30 mai 2012, à l’âge de 94 ans.
Andrew Fielding Huxley, OM 1983; Kt 1974; FRS 1955; PRS 1980–1985, MA, Hon. ScD Cantab. Physiologiste et biophysicien. Né à Hampstead, Londres, le 22 novembre 1917. Démonstrateur, 1946–50, directeur adjoint de la recherche, 1951–59, et lecteur en biophysique expérimentale, 1959–60, Physiological Laboratory, Cambridge ; directeur des études, Trinity College, Cambridge, 1952-1960 ; Professeur Jodrell 1960–69, professeur de recherche de la Royal Society, 1969–83, University College London. Master, 1984–90, Fellow, 1941–60 et depuis 1990, Trinity College, Cambridge (Hon. Fellow, 1967–90). Décédé le 30 mai 2012, à l’âge de 94 ans.
Sir Andrew Huxley, montré dans son laboratoire àFig. 1, était un géant parmi les physiologistes modernes, pionnier dans les domaines de la conduction nerveuse, de l’activation des muscles squelettiques et de la génération de tension. Il a travaillé avec une originalité exceptionnelle et élégante et avait une approche typiquement quantitative de l’analyse physiologique dans un cadre physiquement rigoureux, une approche qui commence maintenant à s’infiltrer de la physiologie et de la biophysique dans leurs sciences biologiques et biomédicales apparentées.
Il est né à Hampstead, Londres, de l’écrivain Leonard Huxley et de Rosalind Bruce en 1917, au sein d’une illustre famille. Son grand-père Thomas Huxley était un biologiste distingué du XIXe siècle et un des premiers partisans de la théorie de l’évolution. Julian Huxley, un pionnier du comportement animal, et Aldous Huxley, l’auteur de Brave New Worldentre autres œuvres, étaient des demi-frères du premier mariage de son père. Il a fait ses études à l’University College (1925–30) et à la Westminster Schools (1930–5), où il s’est inspiré de l’enseignement de JF Rudwick pour passer des classiques aux sciences physiques. Il a choisi de postuler au Trinity College de Cambridge , grâce à l’amitié de sa famille avec George Trevelyan , où il a remporté une importante bourse d’entrée (1935). Ses intérêts se sont finalement tournés vers la physiologie grâce à ses contacts avec Delisle Burns, puis avec ED Adrian, Jack Roughton, William Rushton, Alan Hodgkin et Glenn Millikan, entre autres. À cette fin, ses études se sont poursuivies dans une direction médicale poursuivant l’anatomie en 1937–8 et la physiologie dans la partie II des tripos des sciences naturelles à Cambridge en 1938–39.
 Ces compétences se sont avérées inestimables dans ses travaux scientifiques ultérieurs, lui permettant de concevoir une grande partie de son équipement expérimental. Au cours de sa carrière, il a mis au point un microscope interférentiel pour l’étude des stries dans les fibres musculaires isolées, un microtome pour la réalisation de coupes au microscope électronique et un micromanipulateur, témoignage de l’importance des compétences techniques et des bonnes installations de l’atelier mécanique dont disposait alors physiologistes ( où il a continué à vivre après sa retraite. Cela l’avait amené à concevoir et à construire des microscopes et d’autres instruments scientifiques. Ces compétences se sont avérées inestimables dans ses travaux scientifiques ultérieurs, lui permettant de concevoir une grande partie de son équipement expérimental. Au cours de sa carrière, il a mis au point un microscope interférentiel pour l’étude des stries dans les fibres musculaires isolées, un microtome pour la réalisation de coupes au microscope électronique et un micromanipulateur, témoignage de l’importance des compétences techniques et des bonnes installations de l’atelier mécanique dont disposait alors physiologistes ( où il a continué à vivre après sa retraite. Cela l’avait amené à concevoir et à construire des microscopes et d’autres instruments scientifiques. Ces compétences se sont avérées inestimables dans ses travaux scientifiques ultérieurs, lui permettant de concevoir une grande partie de son équipement expérimental. Au cours de sa carrière, il a mis au point un microscope interférentiel pour l’étude des stries dans les fibres musculaires isolées, un microtome pour la réalisation de coupes au microscope électronique et un micromanipulateur, témoignage de l’importance des compétences techniques et des bonnes installations de l’atelier mécanique dont disposait alors physiologistes.
Ces compétences se sont avérées inestimables dans ses travaux scientifiques ultérieurs, lui permettant de concevoir une grande partie de son équipement expérimental. Au cours de sa carrière, il a mis au point un microscope interférentiel pour l’étude des stries dans les fibres musculaires isolées, un microtome pour la réalisation de coupes au microscope électronique et un micromanipulateur, témoignage de l’importance des compétences techniques et des bonnes installations de l’atelier mécanique dont disposait alors physiologistes ( où il a continué à vivre après sa retraite. Cela l’avait amené à concevoir et à construire des microscopes et d’autres instruments scientifiques. Ces compétences se sont avérées inestimables dans ses travaux scientifiques ultérieurs, lui permettant de concevoir une grande partie de son équipement expérimental. Au cours de sa carrière, il a mis au point un microscope interférentiel pour l’étude des stries dans les fibres musculaires isolées, un microtome pour la réalisation de coupes au microscope électronique et un micromanipulateur, témoignage de l’importance des compétences techniques et des bonnes installations de l’atelier mécanique dont disposait alors physiologistes ( où il a continué à vivre après sa retraite. Cela l’avait amené à concevoir et à construire des microscopes et d’autres instruments scientifiques. Ces compétences se sont avérées inestimables dans ses travaux scientifiques ultérieurs, lui permettant de concevoir une grande partie de son équipement expérimental. Au cours de sa carrière, il a mis au point un microscope interférentiel pour l’étude des stries dans les fibres musculaires isolées, un microtome pour la réalisation de coupes au microscope électronique et un micromanipulateur, témoignage de l’importance des compétences techniques et des bonnes installations de l’atelier mécanique dont disposait alors physiologistes.
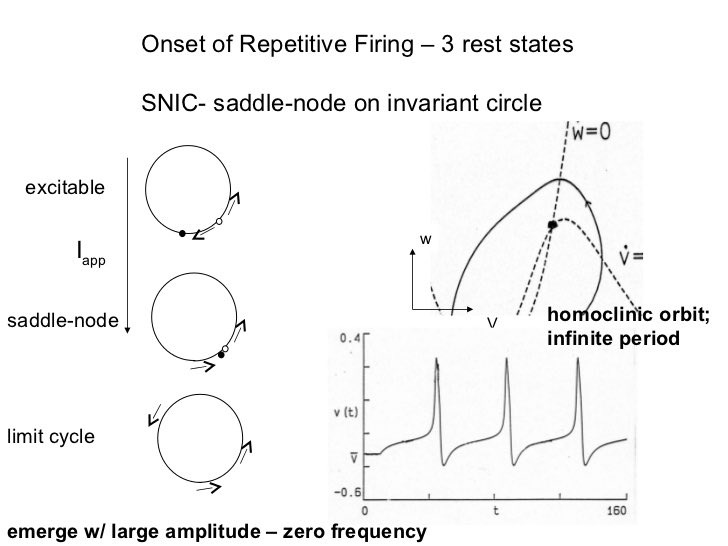
 Le comportement des conductances de sodium et de potassium sous-jacentes pourrait alors être calculé à l’aide de leurs concentrations intracellulaires et extracellulaires connues qui ont fourni les forces motrices de ces courants ( Hodgkin & Huxley, 1952 a , b ). Celles-ci ressemblaient à des transitions de premier ordre incorporant des constantes de vitesse avant et arrière fortement dépendantes de la tension, dont les variables étaient portées respectivement à leurs troisième ( m 3 ) et quatrième ( n 4 ) puissances. La conductance Na + a montré une variable supplémentaire d’inactivation de premier ordre ( h ) qui devait s’avérer d’une importance fondamentale dans la compréhension de la réfractaire post-excitation (Hodgkin & Huxley, 1952c ) .
Le comportement des conductances de sodium et de potassium sous-jacentes pourrait alors être calculé à l’aide de leurs concentrations intracellulaires et extracellulaires connues qui ont fourni les forces motrices de ces courants ( Hodgkin & Huxley, 1952 a , b ). Celles-ci ressemblaient à des transitions de premier ordre incorporant des constantes de vitesse avant et arrière fortement dépendantes de la tension, dont les variables étaient portées respectivement à leurs troisième ( m 3 ) et quatrième ( n 4 ) puissances. La conductance Na + a montré une variable supplémentaire d’inactivation de premier ordre ( h ) qui devait s’avérer d’une importance fondamentale dans la compréhension de la réfractaire post-excitation (Hodgkin & Huxley, 1952c ) .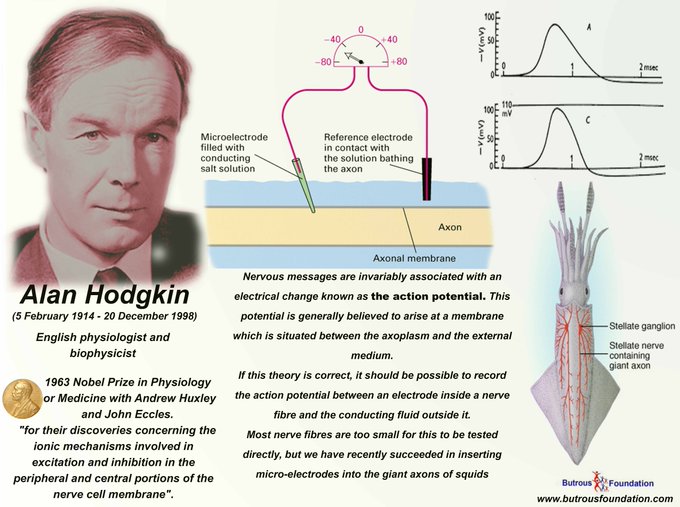
 La dépolarisation résultante produite par le mouvement net vers l’intérieur du Na + résultant vers le bas de son gradient de concentration augmente ensuite davantage la perméabilité au Na + , initiant un processus de rétroaction positive accéléré. Cela ne se termine que lorsque le potentiel de membrane se rapproche du potentiel de Nernst ENa , défini par les concentrations extracellulaires plus élevées de Na + par rapport aux concentrations intracellulaires, lorsque la force motrice nette vers l’intérieur sur Na + devient nulle, et avec le développement plus lent d’une inactivation dépendante de la tension similaire. Une activation similaire, mais plus graduelle, de la perméabilité au K + permettant un mouvement vers l’extérieur du K + le long d’un gradient de concentration opposé contribue également à la restauration du potentiel membranaire à son niveau d’origine. Cela met fin au potentiel d’action, mais un intervalle de temps supplémentaire, la période réfractaire, est nécessaire pour que la perméabilité au Na + retrouve sa capacité d’excitation supplémentaire.
La dépolarisation résultante produite par le mouvement net vers l’intérieur du Na + résultant vers le bas de son gradient de concentration augmente ensuite davantage la perméabilité au Na + , initiant un processus de rétroaction positive accéléré. Cela ne se termine que lorsque le potentiel de membrane se rapproche du potentiel de Nernst ENa , défini par les concentrations extracellulaires plus élevées de Na + par rapport aux concentrations intracellulaires, lorsque la force motrice nette vers l’intérieur sur Na + devient nulle, et avec le développement plus lent d’une inactivation dépendante de la tension similaire. Une activation similaire, mais plus graduelle, de la perméabilité au K + permettant un mouvement vers l’extérieur du K + le long d’un gradient de concentration opposé contribue également à la restauration du potentiel membranaire à son niveau d’origine. Cela met fin au potentiel d’action, mais un intervalle de temps supplémentaire, la période réfractaire, est nécessaire pour que la perméabilité au Na + retrouve sa capacité d’excitation supplémentaire.
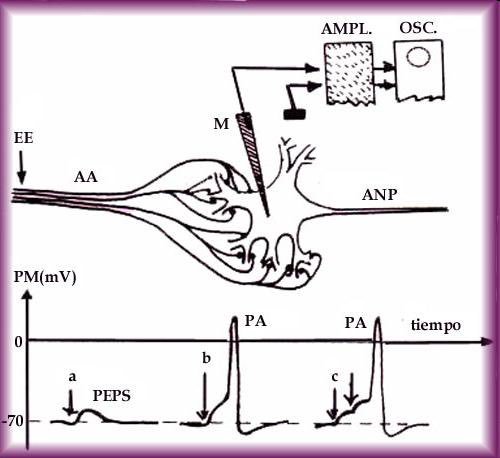

 Ces dernières structures, y compris leur fragilité au changement de volume induit osmotiquement, devaient ensuite être étudiés en détail par, entre autres, Clara Franzini-Armstrong, Robert Eisenberg et Lee Peachey, qui avaient tous commencé leurs travaux scientifiques dans le laboratoire de Huxley. Cela a conduit à l’application de représentations de câbles électriques de leur géométrie tubulaire en utilisant certaines des formulations mathématiques originales de Hodgkin & Huxley pour déterminer leur rôle dans la conduction active de l’excitation à l’intérieur de la fibre ( Adrian & Peachey, 1973 ; Huang & Peachey , 1992 ) . Cela a en outre conduit à la vision récente d’une propagation rapide du potentiel d’action de surface le long de la longueur de la fibre musculaire avec seulement ses composants de basse fréquence filtrés, activant ainsi la capacité tubulaire étendue sans compromettre la conduction de surface. Enfin, les clarifications de l’initiation résultante du couplage excitation-contraction par la détection de tension médiée par le récepteur de dihydropyridine et les processus de libération de calcium couplés allostériquement, médiés par le récepteur de ryanodine ont également utilisé des techniques de blocage de tension. Certaines de ces expériences ont été réalisées en collaboration avec Lee Peachey, qui est revenu à Cambridge à la fin des années 1980 en tant qu’invité de Huxley dans la Master’s Lodge du Trinity College .
Ces dernières structures, y compris leur fragilité au changement de volume induit osmotiquement, devaient ensuite être étudiés en détail par, entre autres, Clara Franzini-Armstrong, Robert Eisenberg et Lee Peachey, qui avaient tous commencé leurs travaux scientifiques dans le laboratoire de Huxley. Cela a conduit à l’application de représentations de câbles électriques de leur géométrie tubulaire en utilisant certaines des formulations mathématiques originales de Hodgkin & Huxley pour déterminer leur rôle dans la conduction active de l’excitation à l’intérieur de la fibre ( Adrian & Peachey, 1973 ; Huang & Peachey , 1992 ) . Cela a en outre conduit à la vision récente d’une propagation rapide du potentiel d’action de surface le long de la longueur de la fibre musculaire avec seulement ses composants de basse fréquence filtrés, activant ainsi la capacité tubulaire étendue sans compromettre la conduction de surface. Enfin, les clarifications de l’initiation résultante du couplage excitation-contraction par la détection de tension médiée par le récepteur de dihydropyridine et les processus de libération de calcium couplés allostériquement, médiés par le récepteur de ryanodine ont également utilisé des techniques de blocage de tension. Certaines de ces expériences ont été réalisées en collaboration avec Lee Peachey, qui est revenu à Cambridge à la fin des années 1980 en tant qu’invité de Huxley dans la Master’s Lodge du Trinity College . Son attribution du prix Nobel de 1963, avec Sir Alan Hodgkin en reconnaissance de leur hypothèse ionique, et Sir John Eccles pour ses travaux sur la signalisation synaptique, ont reconnu ces contributions comme fournissant la base conceptuelle de l’étude de la signalisation cellulaire excitable. Ils ont également suscité une cascade de découvertes importantes avec des implications allant des principes fondamentaux de la fonction des canaux à leur traduction dans les mécanismes de base de la maladie. Le prix Nobel de 1991 devait ensuite être décerné à Erwin Neher et Bert Sakmann pour la nouvelle démonstration directe patch-clamp de petits courants électriques de Na + démontrant directement des événements impliquant des canaux ioniques uniques sous-jacents aux conductances observées . Une adaptation en patch lâche de cette technique a permis de caractériser les distributions des canaux sur la zone membranaire. La dépendance à la tension de la conductance du sodium implique un mécanisme de déclenchement impliquant des mouvements de charge dans le champ électrique transmembranaire. Ceux-ci ont ensuite été démontrés directement sous la forme des courants de déclenchement qui se sont avérés inestimables pour étudier les mécanismes des changements de configuration moléculaire sous-jacents à l’activation des canaux. Cette ligne de travail a récemment abouti à la caractérisation structurale complète du Na+ canal et ses transitions de déclenchement.
Son attribution du prix Nobel de 1963, avec Sir Alan Hodgkin en reconnaissance de leur hypothèse ionique, et Sir John Eccles pour ses travaux sur la signalisation synaptique, ont reconnu ces contributions comme fournissant la base conceptuelle de l’étude de la signalisation cellulaire excitable. Ils ont également suscité une cascade de découvertes importantes avec des implications allant des principes fondamentaux de la fonction des canaux à leur traduction dans les mécanismes de base de la maladie. Le prix Nobel de 1991 devait ensuite être décerné à Erwin Neher et Bert Sakmann pour la nouvelle démonstration directe patch-clamp de petits courants électriques de Na + démontrant directement des événements impliquant des canaux ioniques uniques sous-jacents aux conductances observées . Une adaptation en patch lâche de cette technique a permis de caractériser les distributions des canaux sur la zone membranaire. La dépendance à la tension de la conductance du sodium implique un mécanisme de déclenchement impliquant des mouvements de charge dans le champ électrique transmembranaire. Ceux-ci ont ensuite été démontrés directement sous la forme des courants de déclenchement qui se sont avérés inestimables pour étudier les mécanismes des changements de configuration moléculaire sous-jacents à l’activation des canaux. Cette ligne de travail a récemment abouti à la caractérisation structurale complète du Na+ canal et ses transitions de déclenchement.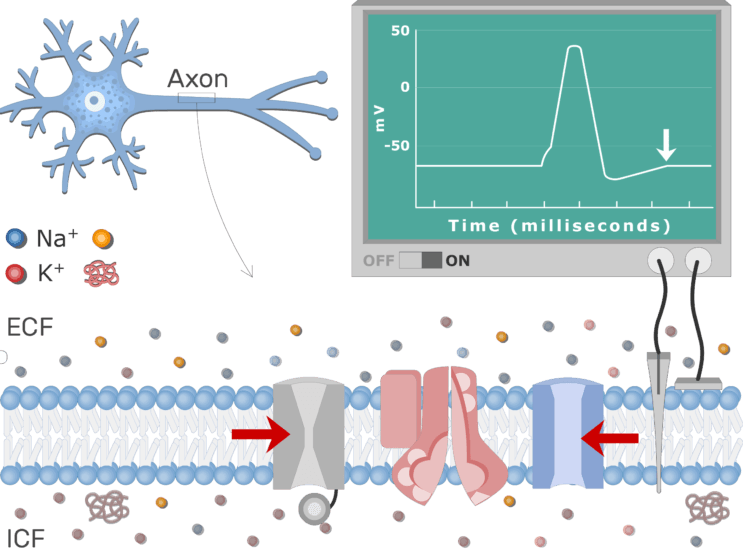 Les techniques de voltage clamp et leurs formulations mathématiques associées ont également été appliquées, corrigées de leur géométrie cellulaire par la théorie des câbles, à d’autres tissus excitables, notamment le nerf myélinisé et le squelette et le muscle cardiaque. Les idées fondamentales ont eu des implications translationnelles larges et fondamentales pour la médecine clinique, y compris la compréhension de l’anesthésie locale et de la douleur, la condition neurologique myotonie congénitale dans le muscle squelettique et l’arythmie dans le muscle cardiaque. Même lorsqu’il avait cessé de travailler activement en laboratoire, Huxley lui-même a encouragé les études récentes de modèles cardiaques murins génétiquement modifiés pour les canalopathies Na + à Cambridge. Ceux-ci ont démontré l’importance de leurs propriétés d’activation et de récupération des canaux Na + par conséquent altérées dans la production d’un stimulateur cardiaque sino-auriculaire et de troubles arythmiques auriculaires et ventriculaires, résultats applicables aux syndromes arythmogènes humains de Brugada et du QT3 long , avec des implications pour la prise en charge des patients souffrant de dysfonctionnement du nœud sinusal, de fibrillation auriculaire et à risque de mort cardiaque subite. Enfin, les formulations de la théorie des circuits électriques décrivant l’hypothèse ionique ont suscité des simulations mathématiques réalistes ultérieures des effets sur l’homéostasie cellulaire des flux non seulement électrogéniques mais aussi électroneutres et osmotiques.
Les techniques de voltage clamp et leurs formulations mathématiques associées ont également été appliquées, corrigées de leur géométrie cellulaire par la théorie des câbles, à d’autres tissus excitables, notamment le nerf myélinisé et le squelette et le muscle cardiaque. Les idées fondamentales ont eu des implications translationnelles larges et fondamentales pour la médecine clinique, y compris la compréhension de l’anesthésie locale et de la douleur, la condition neurologique myotonie congénitale dans le muscle squelettique et l’arythmie dans le muscle cardiaque. Même lorsqu’il avait cessé de travailler activement en laboratoire, Huxley lui-même a encouragé les études récentes de modèles cardiaques murins génétiquement modifiés pour les canalopathies Na + à Cambridge. Ceux-ci ont démontré l’importance de leurs propriétés d’activation et de récupération des canaux Na + par conséquent altérées dans la production d’un stimulateur cardiaque sino-auriculaire et de troubles arythmiques auriculaires et ventriculaires, résultats applicables aux syndromes arythmogènes humains de Brugada et du QT3 long , avec des implications pour la prise en charge des patients souffrant de dysfonctionnement du nœud sinusal, de fibrillation auriculaire et à risque de mort cardiaque subite. Enfin, les formulations de la théorie des circuits électriques décrivant l’hypothèse ionique ont suscité des simulations mathématiques réalistes ultérieures des effets sur l’homéostasie cellulaire des flux non seulement électrogéniques mais aussi électroneutres et osmotiques.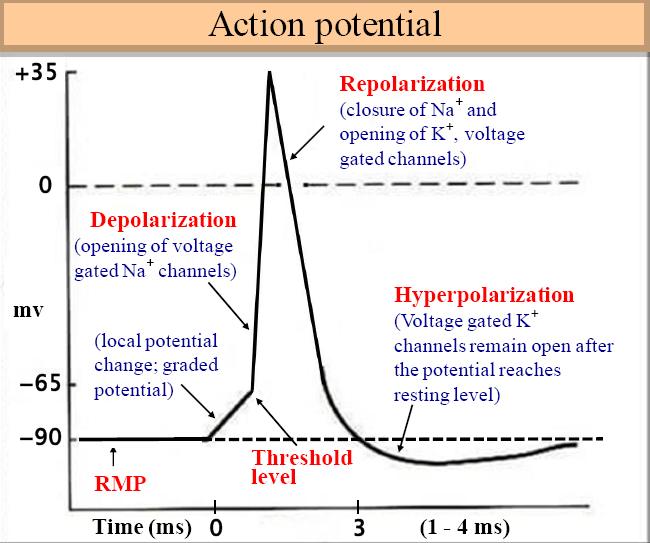 En 1952, Andrew s’est tourné vers la physiologie musculaire. Invité par des observations antérieures au microscope électronique, il a démontré que l’activation électrique de surface initiait une activité mécanique visible en microscopie interférentielle suite à une stimulation localisée à la micropipette uniquement dans des zones spécifiques le long du sarcomère de la fibre musculaire . Celles-ci correspondaient à la ligne Z chez la grenouille couturier mais à la limite entre les bandes A et I dans les muscles du crabe, impliquant ainsi leurs systèmes membranaires tubulaires transversaux invaginants dans l’activation contractile. Ces dernières structures, y compris leur fragilité au changement de volume induit osmotiquement , devaient ensuite être étudiés en détail par, entre autres, Clara Franzini-Armstrong, Robert Eisenberg et Lee Peachey, qui avaient tous commencé leurs travaux scientifiques dans le laboratoire de Huxley.
En 1952, Andrew s’est tourné vers la physiologie musculaire. Invité par des observations antérieures au microscope électronique, il a démontré que l’activation électrique de surface initiait une activité mécanique visible en microscopie interférentielle suite à une stimulation localisée à la micropipette uniquement dans des zones spécifiques le long du sarcomère de la fibre musculaire . Celles-ci correspondaient à la ligne Z chez la grenouille couturier mais à la limite entre les bandes A et I dans les muscles du crabe, impliquant ainsi leurs systèmes membranaires tubulaires transversaux invaginants dans l’activation contractile. Ces dernières structures, y compris leur fragilité au changement de volume induit osmotiquement , devaient ensuite être étudiés en détail par, entre autres, Clara Franzini-Armstrong, Robert Eisenberg et Lee Peachey, qui avaient tous commencé leurs travaux scientifiques dans le laboratoire de Huxley.  Cela a conduit à l’application de représentations de câbles électriques de leur géométrie tubulaire en utilisant certaines des formulations mathématiques originales de Hodgkin & Huxley pour déterminer leur rôle dans la conduction active de l’excitation à l’intérieur de la fibre ( Adrian & Peachey, 1973 ; Huang & Peachey , 1992 ) . Cela a en outre conduit à la vision récente d’une propagation rapide du potentiel d’action de surface le long de la longueur de la fibre musculaire avec seulement ses composants de basse fréquence filtrés, activant ainsi la capacité tubulaire étendue sans compromettre la conduction de surface. Enfin, les clarifications de l’initiation résultante du couplage excitation-contraction par la détection de tension médiée par le récepteur de dihydropyridine et les processus de libération de calcium couplés allostériquement, médiés par le récepteur de ryanodine ont également utilisé des techniques de blocage de tension . Certaines de ces expériences ont été réalisées en collaboration avec Lee Peachey, qui est revenu à Cambridge à la fin des années 1980 en tant qu’invité de Huxley dans la Master’s Lodge du Trinity College .
Cela a conduit à l’application de représentations de câbles électriques de leur géométrie tubulaire en utilisant certaines des formulations mathématiques originales de Hodgkin & Huxley pour déterminer leur rôle dans la conduction active de l’excitation à l’intérieur de la fibre ( Adrian & Peachey, 1973 ; Huang & Peachey , 1992 ) . Cela a en outre conduit à la vision récente d’une propagation rapide du potentiel d’action de surface le long de la longueur de la fibre musculaire avec seulement ses composants de basse fréquence filtrés, activant ainsi la capacité tubulaire étendue sans compromettre la conduction de surface. Enfin, les clarifications de l’initiation résultante du couplage excitation-contraction par la détection de tension médiée par le récepteur de dihydropyridine et les processus de libération de calcium couplés allostériquement, médiés par le récepteur de ryanodine ont également utilisé des techniques de blocage de tension . Certaines de ces expériences ont été réalisées en collaboration avec Lee Peachey, qui est revenu à Cambridge à la fin des années 1980 en tant qu’invité de Huxley dans la Master’s Lodge du Trinity College . Pour ajouter à ces contributions scientifiques capitales, Andrew a été généreux de son temps dans les activités de la Physiological Society, à laquelle il a été élu membre ordinaire en 1942 et membre honoraire en 1979. Il a siégé au comité de rédaction du Journal of Physiology (1950–57) et son Comité (1957–61 ; 1970–74). Il a été coprésident de l’Union internationale des sciences physiologiques de 1986 à 1993. Huxley a également été rédacteur en chef du Journal of Molecular Biology . Il est devenu membre de la Royal Society en 1955 et a siégé à son conseil (1960-1962). Il a travaillé à Woods Hole, Massachusetts, en 1953 en tant que boursier Lalor et a donné les conférences Herter à la Johns Hopkins Medical School (1959); et les conférences Jesup à l’Université de Columbia (1964).
Pour ajouter à ces contributions scientifiques capitales, Andrew a été généreux de son temps dans les activités de la Physiological Society, à laquelle il a été élu membre ordinaire en 1942 et membre honoraire en 1979. Il a siégé au comité de rédaction du Journal of Physiology (1950–57) et son Comité (1957–61 ; 1970–74). Il a été coprésident de l’Union internationale des sciences physiologiques de 1986 à 1993. Huxley a également été rédacteur en chef du Journal of Molecular Biology . Il est devenu membre de la Royal Society en 1955 et a siégé à son conseil (1960-1962). Il a travaillé à Woods Hole, Massachusetts, en 1953 en tant que boursier Lalor et a donné les conférences Herter à la Johns Hopkins Medical School (1959); et les conférences Jesup à l’Université de Columbia (1964).
En 1947, Andrew Huxley épousa Jocelyn Richenda Gammell Pease, fille du généticien MS Pease, et l’hon. HB Pease (née Wedgwood). Elle était juge de paix et était active dans divers travaux publics dans le Cambridgeshire, mais elle est décédée avant lui en 2003. À la fin de ce qui a dû être des journées bien remplies pour eux deux, ils ont pris plaisir à se lire dans le soir, appréciant grandement les œuvres de Jane Austen et laissant la télévision aux petits-enfants les jours de pluie. Ils ont cinq filles et un fils.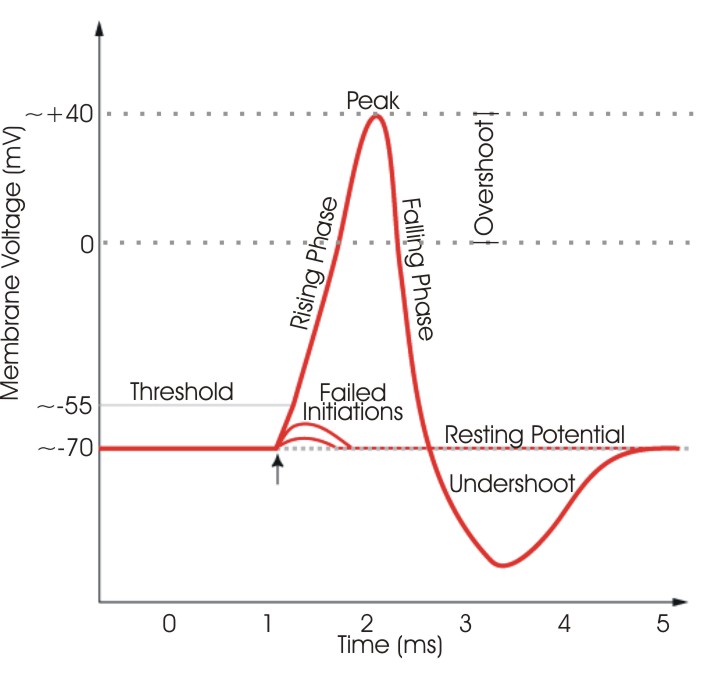 Prix Nobel : Les mesures expérimentales sur lesquelles la paire a basé sa théorie du potentiel d’action représentent l’une des premières applications d’une technique d’ électrophysiologie connue sous le nom de voltage clamp . Le deuxième élément critique de leurs recherches était l’ axone dit géant du calmar de l’Atlantique, qui leur a permis d’enregistrer des courants ioniques comme ils n’auraient pas pu le faire dans presque n’importe quel autre neurone , ces cellules étant trop petites. à étudier par les techniques de l’époque. Les expériences ont eu lieu à l’Université de Cambridge à partir de 1935 avec le nerf sciatique de grenouille et se poursuivant dans les années 1940, après l’interruption de la Seconde Guerre mondiale. Le couple a publié sa théorie en 1952. Dans l’article, ils décrivent l’un des premiers modèles informatiques en biochimie, qui est à la base de la plupart des modèles utilisés en neurobiologie au cours des quatre décennies suivantes. Il a continué à occuper des postes collégiaux et universitaires à Cambridge jusqu’en 1960, date à laquelle il est devenu chef du département de physiologie de l’University College London. En 1969, il a été nommé professeur de recherche de la Royal Society qu’il occupe au département de physiologie de l’University College de Londres. Il maintient actuellement son poste de boursier au Trinity College de Cambridge, où il enseigne la physiologie , les sciences naturelles et la médecine .
Prix Nobel : Les mesures expérimentales sur lesquelles la paire a basé sa théorie du potentiel d’action représentent l’une des premières applications d’une technique d’ électrophysiologie connue sous le nom de voltage clamp . Le deuxième élément critique de leurs recherches était l’ axone dit géant du calmar de l’Atlantique, qui leur a permis d’enregistrer des courants ioniques comme ils n’auraient pas pu le faire dans presque n’importe quel autre neurone , ces cellules étant trop petites. à étudier par les techniques de l’époque. Les expériences ont eu lieu à l’Université de Cambridge à partir de 1935 avec le nerf sciatique de grenouille et se poursuivant dans les années 1940, après l’interruption de la Seconde Guerre mondiale. Le couple a publié sa théorie en 1952. Dans l’article, ils décrivent l’un des premiers modèles informatiques en biochimie, qui est à la base de la plupart des modèles utilisés en neurobiologie au cours des quatre décennies suivantes. Il a continué à occuper des postes collégiaux et universitaires à Cambridge jusqu’en 1960, date à laquelle il est devenu chef du département de physiologie de l’University College London. En 1969, il a été nommé professeur de recherche de la Royal Society qu’il occupe au département de physiologie de l’University College de Londres. Il maintient actuellement son poste de boursier au Trinity College de Cambridge, où il enseigne la physiologie , les sciences naturelles et la médecine .
Sir Andrew est sans doute l’un des plus grands biologistes mathématiciens du XXe siècle. À partir de son travail expérimental avec Hodgkin, il a développé un ensemble d’équations différentielles qui ont fourni une explication mathématique des impulsions nerveuses – le « potentiel d’action ». Ce travail a servi de base à tous les travaux en cours sur les canaux membranaires sensibles au voltage, qui sont responsables du fonctionnement des systèmes nerveux des animaux. Tout à fait séparément, il a développé les équations mathématiques pour le fonctionnement des « ponts croisés » de myosine qui génèrent les forces de glissement entre les filaments d’actine et de myosine, qui provoquent la contraction des muscles squelettiques. Ces équations ont présenté un tout nouveau paradigme pour comprendre la contraction musculaire. En six ans, Huxley et Hodgkin, construisant leur propre équipement, avaient jeté les bases détaillées de la compréhension moderne de la transmission de l’influx nerveux. Ils ont montré que ceux-ci se déplacent, non pas le long du cœur de la fibre, mais le long de la membrane externe sous la forme de cascades successives de deux types d’ions. La découverte et la théorie mathématique détaillée qui accompagnaient les travaux étaient révolutionnaires et leur ont valu leur part du prix Nobel.
En six ans, Huxley et Hodgkin, construisant leur propre équipement, avaient jeté les bases détaillées de la compréhension moderne de la transmission de l’influx nerveux. Ils ont montré que ceux-ci se déplacent, non pas le long du cœur de la fibre, mais le long de la membrane externe sous la forme de cascades successives de deux types d’ions. La découverte et la théorie mathématique détaillée qui accompagnaient les travaux étaient révolutionnaires et leur ont valu leur part du prix Nobel.
Lorsque ce travail a été achevé en 1952, Huxley enseignait la physiologie à Cambridge. Entre autres choses, il s’était intéressé à un phénomène difficile et, à l’époque, inexpliqué : comment le muscle se contracte-t-il ? Pour faire avancer la recherche, il était nécessaire de trouver des moyens d’observer exactement comment le réseau de filaments des fibres musculaires individuelles se comportait pendant la contraction, un problème qui, selon Huxley, appelait un nouveau type de microscope. C’était un défi auquel il ne pouvait résister. Il se trouve que, dans les années qui suivirent immédiatement la guerre, il avait effectué un travail de conception préliminaire sur une forme de microscopie interférentielle qu’il pensait initialement entièrement nouvelle. Il s’est avéré que non seulement d’autres pensaient dans le même sens, mais que des propositions pour des types d’instruments similaires avaient été faites un demi-siècle plus tôt. Huxley a souvent utilisé cet exemple pour montrer à quelle vitesse des idées scientifiques et technologiques précieuses peuvent être oubliées si elles sont incomplètes ou inexploitées.
Il se trouve que, dans les années qui suivirent immédiatement la guerre, il avait effectué un travail de conception préliminaire sur une forme de microscopie interférentielle qu’il pensait initialement entièrement nouvelle. Il s’est avéré que non seulement d’autres pensaient dans le même sens, mais que des propositions pour des types d’instruments similaires avaient été faites un demi-siècle plus tôt. Huxley a souvent utilisé cet exemple pour montrer à quelle vitesse des idées scientifiques et technologiques précieuses peuvent être oubliées si elles sont incomplètes ou inexploitées.
Le développement que Huxley avait en tête permettrait de distinguer les différentes parties d’une seule fibre musculaire striée beaucoup plus facilement et avec une plus grande précision que ce qui peut être atteint par la microscopie conventionnelle. Il était, a-t-il découvert, en avance sur le terrain. Il a donc conçu et construit ses propres microscopes et les a utilisés pour étudier le mécanisme de la contraction et développer en 1954 la théorie du « filament glissant ». (Par coïncidence, son homonyme plus jeune et contemporain, Hugh E Huxley, qui n’était pas apparenté, est arrivé à des conclusions similaires à peu près au même moment).
Né à Hampstead, Huxley est allé à l’école University College puis à Westminster, où il s’est inspiré de l’enseignement des sciences, en particulier de la physique sous JF Rudwick. À Trinity, il s’est impliqué dans le Natural Science Club, qui comptait parmi ses membres plusieurs futurs lauréats du prix Nobel. En tant qu’étudiant de premier cycle, Huxley a trouvé l’inspiration dans un contact continu avec la recherche de troisième cycle et a ensuite travaillé pour empêcher que la séparation ne soit imposée par la création d’instituts de troisième cycle.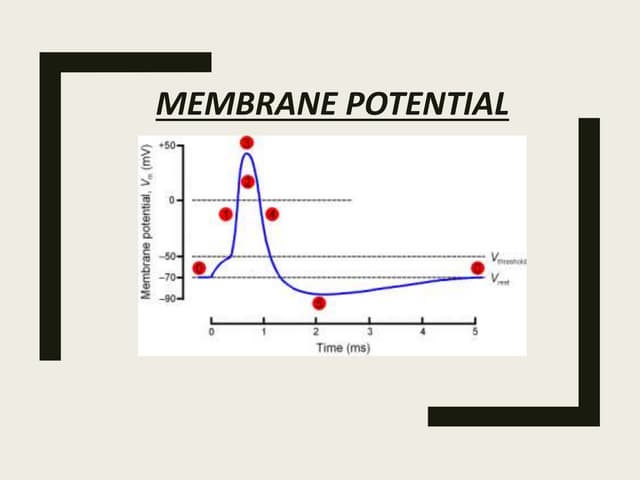 Directeur d’études à Trinity de 1952 à 1960, il a été professeur Jodrell de physiologie à l’University College de Londres de 1960 à 1969, puis a occupé une chaire de recherche de la Royal Society au département de physiologie de l’UCL de 1969 à 1983. Il a été élu membre de la Royal Society en 1955, fait chevalier en 1974 et fait membre de l’Ordre du mérite en 1983.
Directeur d’études à Trinity de 1952 à 1960, il a été professeur Jodrell de physiologie à l’University College de Londres de 1960 à 1969, puis a occupé une chaire de recherche de la Royal Society au département de physiologie de l’UCL de 1969 à 1983. Il a été élu membre de la Royal Society en 1955, fait chevalier en 1974 et fait membre de l’Ordre du mérite en 1983.
L’équilibre, la force et la prudence de la personnalité douce et pacifique de Huxley sont apparus plus clairement lorsqu’il était président de la Royal Society, de 1980 à 1985, une période pendant laquelle la science en Grande-Bretagne était sous la pression du gouvernement. Il s’exprima prudemment mais vivement sur de nombreuses questions de structure scientifique, du rôle de l’université et de la nécessité d’une stabilité à long terme dans la base de recherche nationale.
Andrew F. Huxley (1917-2012) Andrew Fielding Huxley était un physiologiste anglais qui a partagé (avec Sir Alan Hodgkin et Sir John Carew Eccles) le prix Nobel de physiologie ou médecine en 1963. Il a collaboré avec Alan Hodgkin pour élucider les phénomènes chimiques – le mécanisme de la « pompe à sodium » – par lesquels les impulsions nerveuses sont transmises. Il a également effectué d’importants travaux sur la théorie de la contraction musculaire et a participé au développement du microscope interférentiel et de l’ultramicrotome. Il a été fait chevalier en 1974. Il était le petit-fils du célèbre biologiste T.H. Huxley.
Andrew Fielding Huxley était un physiologiste anglais qui a partagé (avec Sir Alan Hodgkin et Sir John Carew Eccles) le prix Nobel de physiologie ou médecine en 1963. Il a collaboré avec Alan Hodgkin pour élucider les phénomènes chimiques – le mécanisme de la « pompe à sodium » – par lesquels les impulsions nerveuses sont transmises. Il a également effectué d’importants travaux sur la théorie de la contraction musculaire et a participé au développement du microscope interférentiel et de l’ultramicrotome. Il a été fait chevalier en 1974. Il était le petit-fils du célèbre biologiste T.H. Huxley.
https://www.bionity.com/en/encyclopedia/Andrew_Huxley.html
https://www.theguardian.com/science/2012/may/31/sir-andrew-huxley
https://www.nobelprize.org/prizes/medicine/1963/huxley/facts/