 Un Homo du Pléistocène moyen de Nesher Ramla, Israël
Un Homo du Pléistocène moyen de Nesher Ramla, Israël Homo qui ? Une nouvelle espèce humaine mystérieuse a été découverte en Israël
Homo qui ? Une nouvelle espèce humaine mystérieuse a été découverte en Israël

 Notre compréhension de l’origine, de la distribution et de l’évolution des premiers humains et de leurs proches parents a été considérablement affinée par de nouvelles informations récentes. Ajoutant à cette tendance, Hershkovitz et al. ont découvert des preuves d’une population Homo archaïque jusque-là inconnue , les « Nesher Ramla Homo » (voir la Perspective de Mirazon Lahr).
Notre compréhension de l’origine, de la distribution et de l’évolution des premiers humains et de leurs proches parents a été considérablement affinée par de nouvelles informations récentes. Ajoutant à cette tendance, Hershkovitz et al. ont découvert des preuves d’une population Homo archaïque jusque-là inconnue , les « Nesher Ramla Homo » (voir la Perspective de Mirazon Lahr). 
 Abstrait : On a longtemps cru que les Néandertaliens étaient originaires et prospéraient sur le continent européen. Cependant, des études morphologiques et génétiques récentes ont suggéré qu’ils pourraient avoir reçu une contribution génétique d’un groupe non européen encore inconnu. Nous rapportons ici la découverte récente de fossiles archaïques d’Homo du site de Nesher Ramla, en Israël, que nous avons datée il y a 140 000 à 120 000 ans. Des analyses qualitatives et quantitatives complètes des os pariétaux, de la mandibule et de la deuxième molaire inférieure ont révélé que cet Homogroupe présente une combinaison distinctive de caractéristiques néandertaliennes et archaïques. Nous suggérons que ces spécimens représentent les derniers survivants d’un paléodème du Pléistocène moyen du Levant qui a très probablement été impliqué dans l’évolution de l’ Homo du Pléistocène moyen en Europe et en Asie de l’Est.
Abstrait : On a longtemps cru que les Néandertaliens étaient originaires et prospéraient sur le continent européen. Cependant, des études morphologiques et génétiques récentes ont suggéré qu’ils pourraient avoir reçu une contribution génétique d’un groupe non européen encore inconnu. Nous rapportons ici la découverte récente de fossiles archaïques d’Homo du site de Nesher Ramla, en Israël, que nous avons datée il y a 140 000 à 120 000 ans. Des analyses qualitatives et quantitatives complètes des os pariétaux, de la mandibule et de la deuxième molaire inférieure ont révélé que cet Homogroupe présente une combinaison distinctive de caractéristiques néandertaliennes et archaïques. Nous suggérons que ces spécimens représentent les derniers survivants d’un paléodème du Pléistocène moyen du Levant qui a très probablement été impliqué dans l’évolution de l’ Homo du Pléistocène moyen en Europe et en Asie de l’Est.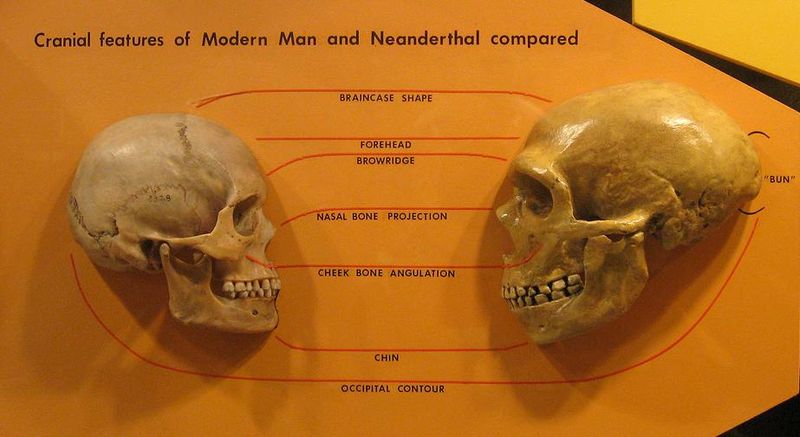 De récentes études dentaires , mandibulaires, génétiques et démographiques ont prédit l’existence d’une population encore non identifiée d’Afrique ou d’Asie occidentale du Pléistocène moyen (MP) qui a contribué à l’évolution du clade néandertalien. Cela contraste avec la vision traditionnelle qui considérait le continent européen comme le seul lieu d’origine des Néandertaliens et de leurs ancêtres directs. Nous rapportons ici la découverte de plusieurs fossiles provenant du site en plein air MP récemment fouillé de Nesher Ramla (NR), au centre d’Israël ( Fig. 1 ), en association avec des artefacts en pierre et des restes fauniques.
De récentes études dentaires , mandibulaires, génétiques et démographiques ont prédit l’existence d’une population encore non identifiée d’Afrique ou d’Asie occidentale du Pléistocène moyen (MP) qui a contribué à l’évolution du clade néandertalien. Cela contraste avec la vision traditionnelle qui considérait le continent européen comme le seul lieu d’origine des Néandertaliens et de leurs ancêtres directs. Nous rapportons ici la découverte de plusieurs fossiles provenant du site en plein air MP récemment fouillé de Nesher Ramla (NR), au centre d’Israël ( Fig. 1 ), en association avec des artefacts en pierre et des restes fauniques.  Fig. 1 Le site de Nesher Ramla et sa stratigraphie. 🙁 A ) Carte de la région de la Méditerranée orientale montrant l’emplacement du site. ( B ) Vue de la fouille depuis l’est. ( C ) Coupe de la séquence archéologique générale insérée dans la géologie locale (l’échelle verticale à gauche indique les mètres au-dessus du niveau de la mer). ( D ) Grossissement des unités IV à VII. Une étoile rouge indique l’emplacement du NR-1 (os pariétal) à 8 m sous la surface dans l’unité VI. NR-2 (mandibule) a été trouvé dans la même unité, à 2 m au nord.
Fig. 1 Le site de Nesher Ramla et sa stratigraphie. 🙁 A ) Carte de la région de la Méditerranée orientale montrant l’emplacement du site. ( B ) Vue de la fouille depuis l’est. ( C ) Coupe de la séquence archéologique générale insérée dans la géologie locale (l’échelle verticale à gauche indique les mètres au-dessus du niveau de la mer). ( D ) Grossissement des unités IV à VII. Une étoile rouge indique l’emplacement du NR-1 (os pariétal) à 8 m sous la surface dans l’unité VI. NR-2 (mandibule) a été trouvé dans la même unité, à 2 m au nord.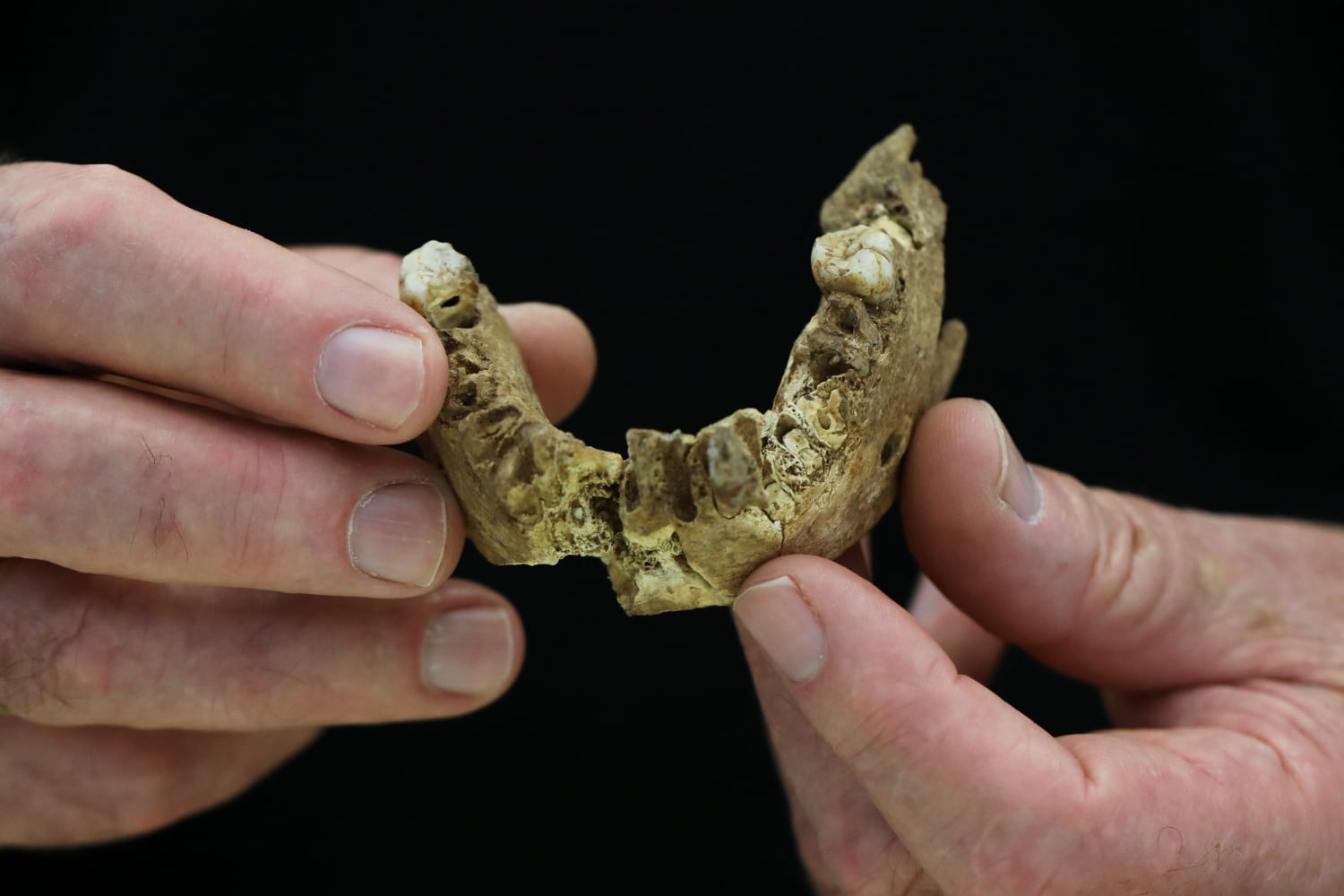 Un os pariétal droit presque complet et quatre fragments de l’os pariétal gauche représentent le fossile NR-1 ( Fig. 2A et fig. S1). Le fossile NR-2 est une mandibule presque complète, ne manquant que la branche gauche, le processus condylien droit et l’angle mandibulaire de la branche droite ( Fig. 3 ). La deuxième molaire inférieure gauche (NR-2 M 2 ) et la plupart des racines dentaires sont toujours en place (fig. S2). NR-1 et NR-2 ont été trouvés in situ dans la couche archéologique la plus basse ( Fig. 1D , Unité VI), avec des os d’animaux et des outils en silex, et représentent très probablement le même individu (texte complémentaire A).
Un os pariétal droit presque complet et quatre fragments de l’os pariétal gauche représentent le fossile NR-1 ( Fig. 2A et fig. S1). Le fossile NR-2 est une mandibule presque complète, ne manquant que la branche gauche, le processus condylien droit et l’angle mandibulaire de la branche droite ( Fig. 3 ). La deuxième molaire inférieure gauche (NR-2 M 2 ) et la plupart des racines dentaires sont toujours en place (fig. S2). NR-1 et NR-2 ont été trouvés in situ dans la couche archéologique la plus basse ( Fig. 1D , Unité VI), avec des os d’animaux et des outils en silex, et représentent très probablement le même individu (texte complémentaire A).  Fig. 2 Les os pariétaux NR-1 et leurs analyses. ( A ) Reconstruction virtuelle de la partie médiane de la calotte crânienne incluant les deux pariétaux. Le pariétal droit presque complet a été reflété en miroir et les fragments pariétaux gauches (représentés en différentes couleurs) ont été superposés, montrant un très bon recouvrement avec la partie controlatérale. ( B ) Un arbre phylogénétique non enraciné construit sur la forme moyenne de chaque groupe en utilisant la méthode de jointure voisine.
Fig. 2 Les os pariétaux NR-1 et leurs analyses. ( A ) Reconstruction virtuelle de la partie médiane de la calotte crânienne incluant les deux pariétaux. Le pariétal droit presque complet a été reflété en miroir et les fragments pariétaux gauches (représentés en différentes couleurs) ont été superposés, montrant un très bon recouvrement avec la partie controlatérale. ( B ) Un arbre phylogénétique non enraciné construit sur la forme moyenne de chaque groupe en utilisant la méthode de jointure voisine.  La NR-1 est positionnée sur la ligne menant au MP Homo africain , proche du point de scission de H. erectus et non loin du MP Homo européen et des Néandertaliens. ( C ) Parcelle PCA dans l’espace de forme pour l’os pariétal. PC1 sépare H. sapiens précoce et récentdu reste de l’ échantillon Homo . Le MP Homo européen et les Néandertaliens (y compris le Levantin Amud 1) se chevauchent et se distinguent de l’ H. erectus asiatique et du MP Homo africain le long de PC3 ; NR-1 est intermédiaire entre ces groupes et est proche de Petralona et de certains H. erectus d’Asie tardive . Les formes extrêmes le long des PC sont représentées à partir des vues antérieure et médiale.
La NR-1 est positionnée sur la ligne menant au MP Homo africain , proche du point de scission de H. erectus et non loin du MP Homo européen et des Néandertaliens. ( C ) Parcelle PCA dans l’espace de forme pour l’os pariétal. PC1 sépare H. sapiens précoce et récentdu reste de l’ échantillon Homo . Le MP Homo européen et les Néandertaliens (y compris le Levantin Amud 1) se chevauchent et se distinguent de l’ H. erectus asiatique et du MP Homo africain le long de PC3 ; NR-1 est intermédiaire entre ces groupes et est proche de Petralona et de certains H. erectus d’Asie tardive . Les formes extrêmes le long des PC sont représentées à partir des vues antérieure et médiale. Fig. 3 La mandibule NR-2 et son analyse.: ( A ) Différentes vues de la mandibule reconstruite. ( B ) Arbre phylogénétique non raciné construit en utilisant la forme moyenne de chaque groupe Homo en utilisant la méthode de jonction des voisins. NR-2 et Tabun C1 ont un ancêtre commun qui est proche de la scission entre les Néandertaliens et le MP Homo européen et loin des premiers et récents H. s apiens , ainsi que de H. erectus et du MP Homo africain . ( C ) Parcelle PCA pour la mandibule dans l’espace de forme. La combinaison de PC1 et PC2 sépare les H. sapiens précoces et récents des autres spécimens d’Homo et distingue les MP européensHomo et Atapuerca des Néandertaliens (dont le Levantin Amud 1) et des H. erectus asiatiques . Parcelles NR-2 entre les Néandertaliens et le député européen Homo. Les formes extrêmes le long de PC1 et PC2 sont représentées à partir d’une vue latérale
Fig. 3 La mandibule NR-2 et son analyse.: ( A ) Différentes vues de la mandibule reconstruite. ( B ) Arbre phylogénétique non raciné construit en utilisant la forme moyenne de chaque groupe Homo en utilisant la méthode de jonction des voisins. NR-2 et Tabun C1 ont un ancêtre commun qui est proche de la scission entre les Néandertaliens et le MP Homo européen et loin des premiers et récents H. s apiens , ainsi que de H. erectus et du MP Homo africain . ( C ) Parcelle PCA pour la mandibule dans l’espace de forme. La combinaison de PC1 et PC2 sépare les H. sapiens précoces et récents des autres spécimens d’Homo et distingue les MP européensHomo et Atapuerca des Néandertaliens (dont le Levantin Amud 1) et des H. erectus asiatiques . Parcelles NR-2 entre les Néandertaliens et le député européen Homo. Les formes extrêmes le long de PC1 et PC2 sont représentées à partir d’une vue latérale
 Les éléments anatomiques préservés ont été minutieusement décrits et analysés par rapport à un grand nombre de fossiles de différentes périodes (tableau S1), en utilisant une combinaison d’approches traditionnelles basées sur des mesures linéaires et angulaires, ainsi que des repères tridimensionnels (3D). méthodes de morphométrie géométrique (GM) (texte complémentaire C à E).
Les éléments anatomiques préservés ont été minutieusement décrits et analysés par rapport à un grand nombre de fossiles de différentes périodes (tableau S1), en utilisant une combinaison d’approches traditionnelles basées sur des mesures linéaires et angulaires, ainsi que des repères tridimensionnels (3D). méthodes de morphométrie géométrique (GM) (texte complémentaire C à E).
La morphologie globale des os pariétaux NR-1 (texte complémentaire C et tableaux S2 et S3) indique une voûte crânienne archaïque et basse, typique des spécimens MP Homo et sensiblement différente des H. sapiens anciens et récents , qui manifestent plutôt un os pariétal incurvé avec une éminence prononcée. Un autre support pour la morphologie plutôt archaïque de l’ Homo NR vient de l’angle formé par les sutures coronales et sagittales (angle c/s), 91,1° dans le spécimen NR-1. Cet angle a augmenté au cours de l’évolution de l’Homo pléistocène (fig. S3) : H. erectus et MP Homo africain présentent un angle moyen de 92,1° ± 2,1° ; l’angle s’ouvre à 94,9° ± 3,4° chez les MP Homo /Néandertaliens européens et atteint 99,4° ± 4,2° chez les H. sapiens anciens et récents . L’angle c/s est significativement différent entre ces trois groupes ( H = 22,5, p < 0,001). L’angle c/s pour NR-1 est similaire à celui de l’Homo archaïque, en particulier African MP Homo (91,1° ± 1,1°), et se situe en dehors de la plage de variation de H. sapiens .
Un autre support pour la morphologie plutôt archaïque de l’ Homo NR vient de l’angle formé par les sutures coronales et sagittales (angle c/s), 91,1° dans le spécimen NR-1. Cet angle a augmenté au cours de l’évolution de l’Homo pléistocène (fig. S3) : H. erectus et MP Homo africain présentent un angle moyen de 92,1° ± 2,1° ; l’angle s’ouvre à 94,9° ± 3,4° chez les MP Homo /Néandertaliens européens et atteint 99,4° ± 4,2° chez les H. sapiens anciens et récents . L’angle c/s est significativement différent entre ces trois groupes ( H = 22,5, p < 0,001). L’angle c/s pour NR-1 est similaire à celui de l’Homo archaïque, en particulier African MP Homo (91,1° ± 1,1°), et se situe en dehors de la plage de variation de H. sapiens . L’os pariétal NR-1 est considérablement épais, principalement dans la zone de l’éminence pariétale (fig. S4 et S5). Sur cet aspect, le pariétal NR-1 est similaire à celui des spécimens MP Homo européens (ex. Petralona, Atapuerca SH) (fig. S4). Il est généralement plus épais que le pariétal des Néandertaliens (par exemple, Amud 1, Guattari et La Chapelle-aux-Saints) et la plupart des premiers H. sapiens (sauf Laetoli H18 et Omo 2), et il est beaucoup plus épais que celui des H. sapiens . L’analyse 3D GM, utilisée pour évaluer la variation de forme de NR-1 par rapport à un échantillon comparatif de Pléistocène et d’Homo récent (texte complémentaire, Fig. 2C , fig. S1 et tableau S1), confirme la morphologie archaïque de NR-1. Les trois premières composantes principales (PC) expliquent 74,5 % de la variance de forme totale. Le premier PC (34,9%) différencie les H. sapiens précoces et récents de tous les autres groupes, y compris les H. erectus asiatiques, les MP Homo européens et africains et les Néandertaliens, en raison de leur courbure marquée le long des plans sagittal et coronal ( Fig. 2C ). Le deuxième PC (21,3 %) n’est pas taxonomiquement informatif (fig. S1).
L’os pariétal NR-1 est considérablement épais, principalement dans la zone de l’éminence pariétale (fig. S4 et S5). Sur cet aspect, le pariétal NR-1 est similaire à celui des spécimens MP Homo européens (ex. Petralona, Atapuerca SH) (fig. S4). Il est généralement plus épais que le pariétal des Néandertaliens (par exemple, Amud 1, Guattari et La Chapelle-aux-Saints) et la plupart des premiers H. sapiens (sauf Laetoli H18 et Omo 2), et il est beaucoup plus épais que celui des H. sapiens . L’analyse 3D GM, utilisée pour évaluer la variation de forme de NR-1 par rapport à un échantillon comparatif de Pléistocène et d’Homo récent (texte complémentaire, Fig. 2C , fig. S1 et tableau S1), confirme la morphologie archaïque de NR-1. Les trois premières composantes principales (PC) expliquent 74,5 % de la variance de forme totale. Le premier PC (34,9%) différencie les H. sapiens précoces et récents de tous les autres groupes, y compris les H. erectus asiatiques, les MP Homo européens et africains et les Néandertaliens, en raison de leur courbure marquée le long des plans sagittal et coronal ( Fig. 2C ). Le deuxième PC (21,3 %) n’est pas taxonomiquement informatif (fig. S1). Le troisième PC (18,3%) sépare les AsiatiquesH. erectus et MP Homo africain des Néandertaliens et MP Homo européen ( Fig. 2C ), sur la base du développement relatif de l’éminence pariétale et de sa position antéro-postérieure relative. Le groupe MP européen est caractérisé par un os pariétal antéro-postérieur et supéro-inférieur plus plat ( Fig. 2C ). NR-1 est distinct de H. sapiens ; il se situe à une position intermédiaire entre les amas de Néandertal et de MP Homo ( Fig. 2C ). Une analyse phylogénétique non enracinée, basée sur la forme moyenne de chaque groupe Homo , a placé NR-1 près de l’origine de la branche menant à l’ homo MP africain., proche de la scission de la branche H. erectus et des Homo MP européens et des Néandertaliens (dont Atapuerca SH), et loin des H. sapiens anciens et récents ( Fig. 2B ).
Le troisième PC (18,3%) sépare les AsiatiquesH. erectus et MP Homo africain des Néandertaliens et MP Homo européen ( Fig. 2C ), sur la base du développement relatif de l’éminence pariétale et de sa position antéro-postérieure relative. Le groupe MP européen est caractérisé par un os pariétal antéro-postérieur et supéro-inférieur plus plat ( Fig. 2C ). NR-1 est distinct de H. sapiens ; il se situe à une position intermédiaire entre les amas de Néandertal et de MP Homo ( Fig. 2C ). Une analyse phylogénétique non enracinée, basée sur la forme moyenne de chaque groupe Homo , a placé NR-1 près de l’origine de la branche menant à l’ homo MP africain., proche de la scission de la branche H. erectus et des Homo MP européens et des Néandertaliens (dont Atapuerca SH), et loin des H. sapiens anciens et récents ( Fig. 2B ). En ce qui concerne la configuration de la surface endopariétale (fig. S5), NR-1 est polygonale, c’est-à-dire que la surface est clairement orientée selon trois plans distincts (fig. S5). Au lieu de cela, les Néandertaliens et H. sapiens manifestent une surface endopariétale arquée. La planéité du lobule pariétal supérieur, vue dans l’endocast virtuel NR-1 (fig. S6), est l’une des caractéristiques les plus caractéristiques de MP Homo . Autres caractéristiques importantes de l’endocast NR-1 et qui sont également typiques de MP Homosont la position très basse de la largeur endocrânienne maximale à la partie supérieure de la première circonvolution temporale (fig. S6), le lobe pariétal très court (fig. S7), ainsi que les longueurs différentes de la largeur endocrânienne maximale et de la largeur intrapariétale ainsi que leur position postérieure sur l’os pariétal (tableau S3). Ces caractéristiques peuvent parfois être également observées chez les Néandertaliens (tableau S3) . Inversement, les spécimens récents d’H. sapiens présentent des largeurs endocrâniennes et intrapariétales maximales inférieures à l’égalité, qui sont situées beaucoup plus haut et plus en avant que dans NR-1.
En ce qui concerne la configuration de la surface endopariétale (fig. S5), NR-1 est polygonale, c’est-à-dire que la surface est clairement orientée selon trois plans distincts (fig. S5). Au lieu de cela, les Néandertaliens et H. sapiens manifestent une surface endopariétale arquée. La planéité du lobule pariétal supérieur, vue dans l’endocast virtuel NR-1 (fig. S6), est l’une des caractéristiques les plus caractéristiques de MP Homo . Autres caractéristiques importantes de l’endocast NR-1 et qui sont également typiques de MP Homosont la position très basse de la largeur endocrânienne maximale à la partie supérieure de la première circonvolution temporale (fig. S6), le lobe pariétal très court (fig. S7), ainsi que les longueurs différentes de la largeur endocrânienne maximale et de la largeur intrapariétale ainsi que leur position postérieure sur l’os pariétal (tableau S3). Ces caractéristiques peuvent parfois être également observées chez les Néandertaliens (tableau S3) . Inversement, les spécimens récents d’H. sapiens présentent des largeurs endocrâniennes et intrapariétales maximales inférieures à l’égalité, qui sont situées beaucoup plus haut et plus en avant que dans NR-1. Le schéma vasculaire des vaisseaux méningés moyens dans NR-1 est simple. Seules quelques courtes ramifications sont visibles et les anastomoses sont absentes, comme c’est le cas chez d’autres MP Homo et Néandertaliens (fig. S9 et S10). La branche postérieure du vaisseau méningé moyen dans NR-1 est aussi développée que la branche antérieure, un schéma persistant chez MP Homo . Les Néandertaliens (par exemple, La Quina H5 et La Chapelle-aux-Saints) et les H. sapiens récents montrent une dominance de la branche antérieure ; ce dernier possède également des empreintes endocrâniennes vasculaires complexes (fig. S10).
Le schéma vasculaire des vaisseaux méningés moyens dans NR-1 est simple. Seules quelques courtes ramifications sont visibles et les anastomoses sont absentes, comme c’est le cas chez d’autres MP Homo et Néandertaliens (fig. S9 et S10). La branche postérieure du vaisseau méningé moyen dans NR-1 est aussi développée que la branche antérieure, un schéma persistant chez MP Homo . Les Néandertaliens (par exemple, La Quina H5 et La Chapelle-aux-Saints) et les H. sapiens récents montrent une dominance de la branche antérieure ; ce dernier possède également des empreintes endocrâniennes vasculaires complexes (fig. S10). Le spécimen NR-2 est une mandibule robuste ( Fig. 3 ) ; le corpus est large médio-latéralement, et l’os cortical est épais ( Fig. 3 , fig. S11). Sa caractéristique la plus prononcée est la branche courte par rapport à la hauteur du corps, avec un processus coronoïde robuste, bas et large ( Fig. 3 ). Ce spécimen présente plusieurs caractéristiques archaïques (par exemple, pas de trigonum mentale ou d’incurvatio mandibulae, une large incisura submentalis, un planum alveolare développé, un planum triangulare fortement développé et un corpus mandibulaire qui présente des marges alvéolaires et basales assez parallèles) couramment observées chez MP Homo (texte complémentaire D et tableau S4A).
Le spécimen NR-2 est une mandibule robuste ( Fig. 3 ) ; le corpus est large médio-latéralement, et l’os cortical est épais ( Fig. 3 , fig. S11). Sa caractéristique la plus prononcée est la branche courte par rapport à la hauteur du corps, avec un processus coronoïde robuste, bas et large ( Fig. 3 ). Ce spécimen présente plusieurs caractéristiques archaïques (par exemple, pas de trigonum mentale ou d’incurvatio mandibulae, une large incisura submentalis, un planum alveolare développé, un planum triangulare fortement développé et un corpus mandibulaire qui présente des marges alvéolaires et basales assez parallèles) couramment observées chez MP Homo (texte complémentaire D et tableau S4A).
Nous avons combiné les caractéristiques mandibulaires pertinentes sur le plan taxonomique dans une analyse de regroupement hiérarchique (fig. S12). Les humains modernes et pléistocènes forment les deux groupes principaux : NR-2 est placé sur une branche latérale de ce dernier, avec MP Homo d’Atapuerca SH, Tighenif 3, Arago XIII et un Néandertalien (fig. S12). Les traits discrets soulignent la nature mosaïque de la mandibule NR, montrant une morphologie archaïque ainsi que certains traits néandertaliens. Les dimensions métriques du corps mandibulaire NR-2 sont présentées sur la fig. S13. La zone symphysaire est considérablement épaisse (16,6 mm), proche des valeurs des mandibules européennes MP Homo (16,9 ± 2,1 mm), et moyennement haute (33,7 mm), proche de la moyenne néandertalienne (34,0 ± 4,6 mm). Le corps (mesuré entre la première et la deuxième molaire) est épais (17,7 mm), dans la fourchette du MP Homo européen (18,1 ± 3,1 mm), mais plus grand (32,7 mm) que celui du MP Homo européen (30,2 ± 1,6 mm ) et Néandertaliens (29,9 ± 3,3 mm), proches des valeurs des premiers H. sapiens (33,0 ± 4,0 mm). Les résultats de l’analyse 3D GM (fig. S14 et tableaux S5A et S5B) pour la mandibule NR-2 sont illustrés sur la Fig. 3C . Les deux premières composantes principales expliquent 47,5 % de la variance totale. La variation le long de PC1 (37,9%) est entraînée par des changements dans la longueur du corps mandibulaire, la forme de la branche (plus courte et plus large chez les Homo archaïques ) et l’expression de la zone mentale. La variation le long de PC2 (9,6%) reflète les changements de hauteur du corps (principalement dans la région mentale) et la transition des marges alvéolaires et basales parallèles d’un corps à celles qui convergent vers l’arrière.
Les dimensions métriques du corps mandibulaire NR-2 sont présentées sur la fig. S13. La zone symphysaire est considérablement épaisse (16,6 mm), proche des valeurs des mandibules européennes MP Homo (16,9 ± 2,1 mm), et moyennement haute (33,7 mm), proche de la moyenne néandertalienne (34,0 ± 4,6 mm). Le corps (mesuré entre la première et la deuxième molaire) est épais (17,7 mm), dans la fourchette du MP Homo européen (18,1 ± 3,1 mm), mais plus grand (32,7 mm) que celui du MP Homo européen (30,2 ± 1,6 mm ) et Néandertaliens (29,9 ± 3,3 mm), proches des valeurs des premiers H. sapiens (33,0 ± 4,0 mm). Les résultats de l’analyse 3D GM (fig. S14 et tableaux S5A et S5B) pour la mandibule NR-2 sont illustrés sur la Fig. 3C . Les deux premières composantes principales expliquent 47,5 % de la variance totale. La variation le long de PC1 (37,9%) est entraînée par des changements dans la longueur du corps mandibulaire, la forme de la branche (plus courte et plus large chez les Homo archaïques ) et l’expression de la zone mentale. La variation le long de PC2 (9,6%) reflète les changements de hauteur du corps (principalement dans la région mentale) et la transition des marges alvéolaires et basales parallèles d’un corps à celles qui convergent vers l’arrière.  Dans la parcelle PC1-PC2, les H. sapiens précoces et récents se séparent des autres spécimens d’Homo , alors que les Homo MP européenset Atapuerca SH se distinguent des Néandertaliens (dont le Levantin Amud 1) et des H. erectus asiatiques . NR-2 se situe entre les Néandertaliens et les spécimens européens MP Homo (y compris Atapuerca SH), bien en dehors de la plage de variation de H. sapiens. L’analyse phylogénétique, basée sur la forme moyenne mandibulaire de chaque groupe d’hominines, a placé NR-2 sur une branche distincte (avec Tabun C1), proche de la scission entre MP fossiles européens et Néandertaliens, et loin de H. erectus, MP africain Homo et H. sapiens ( Fig. 3B). Ce résultat, basé uniquement sur les métriques, fait largement écho aux résultats de l’analyse typologique basée sur des traits discrets et confirme que NR-2 appartient à un groupe archaïque aux affinités néandertaliennes.
Dans la parcelle PC1-PC2, les H. sapiens précoces et récents se séparent des autres spécimens d’Homo , alors que les Homo MP européenset Atapuerca SH se distinguent des Néandertaliens (dont le Levantin Amud 1) et des H. erectus asiatiques . NR-2 se situe entre les Néandertaliens et les spécimens européens MP Homo (y compris Atapuerca SH), bien en dehors de la plage de variation de H. sapiens. L’analyse phylogénétique, basée sur la forme moyenne mandibulaire de chaque groupe d’hominines, a placé NR-2 sur une branche distincte (avec Tabun C1), proche de la scission entre MP fossiles européens et Néandertaliens, et loin de H. erectus, MP africain Homo et H. sapiens ( Fig. 3B). Ce résultat, basé uniquement sur les métriques, fait largement écho aux résultats de l’analyse typologique basée sur des traits discrets et confirme que NR-2 appartient à un groupe archaïque aux affinités néandertaliennes. La deuxième molaire inférieure (NR-2 M 2 ) est complète et présente une certaine usure occlusale provoquant une légère exposition des cornes dentinaires ( Fig. 4A et texte complémentaire E). La surface occlusale du NR-2 M 2 révèle quatre cuspides bien développées et un hypoconulide. La présence de cinq cuspides principales est typique pour la plupart (70%) des Atapuerca SH Homo et des Néandertaliens. Le NR-2 M 2 a une crête médiane trigonide continue claire et une crête trigonide distale discontinue sur la surface de la dentine, correspondant au grade 3 de Bailey et al . (fig. S15). Une crête mi-trigonide est présente chez plus de 90% des Néandertaliens et MP Homo d’Atapuerca SH. Une expression de grade 3 de la crête mi-trigonide, comme dans le NR-2 M 2 , est présente chez près de 60 % des spécimens néandertaliens, mais elle est absente chez H. sapiens. Le spécimen Qesem Cave M 2 (QC-J15) montre un schéma similaire d’une crête continue à mi-trigonide et d’une crête discontinue à trigonide distal (fig. S15). Le spécimen Ehringsdorf G ne présente qu’une crête mi-trigonide (mais pas de crête distale) (fig. S15), alors que le spécimen Mauer ne manifeste pas du tout de crête mi-trigonide.
La deuxième molaire inférieure (NR-2 M 2 ) est complète et présente une certaine usure occlusale provoquant une légère exposition des cornes dentinaires ( Fig. 4A et texte complémentaire E). La surface occlusale du NR-2 M 2 révèle quatre cuspides bien développées et un hypoconulide. La présence de cinq cuspides principales est typique pour la plupart (70%) des Atapuerca SH Homo et des Néandertaliens. Le NR-2 M 2 a une crête médiane trigonide continue claire et une crête trigonide distale discontinue sur la surface de la dentine, correspondant au grade 3 de Bailey et al . (fig. S15). Une crête mi-trigonide est présente chez plus de 90% des Néandertaliens et MP Homo d’Atapuerca SH. Une expression de grade 3 de la crête mi-trigonide, comme dans le NR-2 M 2 , est présente chez près de 60 % des spécimens néandertaliens, mais elle est absente chez H. sapiens. Le spécimen Qesem Cave M 2 (QC-J15) montre un schéma similaire d’une crête continue à mi-trigonide et d’une crête discontinue à trigonide distal (fig. S15). Le spécimen Ehringsdorf G ne présente qu’une crête mi-trigonide (mais pas de crête distale) (fig. S15), alors que le spécimen Mauer ne manifeste pas du tout de crête mi-trigonide.  Fig. 4 La deuxième molaire inférieure gauche (NR2 M 2 ) et son analyse.: ( A ) De gauche, le NR2 M 2 en vue occlusale, avec et sans coiffe en émail, et en vues mésiale, buccale, distale et linguale. Les facettes d’usure interproximales mésiales et distales sont visibles sur les vues vestibulaire et linguale. ( B ) Une construction d’arbre phylogénétique non racinée, basée sur la forme moyenne de chaque groupe d’hominines en utilisant la méthode de jonction des voisins. NR2 M 2 est proche des Néandertaliens et éloigné d’H. sapiens . ( C ) Canaux radiculaires (cavités pulpaires) dans les vues mésiale, buccale, distale et linguale. Les racines pyramidales et la cavité pulpaire taurodontique se prolongent dans l’apex avant de se ramifier en canaux radiculaires courts. ( D) Les racines dentaires (en marron) et leurs canaux (en rouge) sont présentés en images « verre » de la mandibule en vue latérale, révélant la présence de taurodontisme.
Fig. 4 La deuxième molaire inférieure gauche (NR2 M 2 ) et son analyse.: ( A ) De gauche, le NR2 M 2 en vue occlusale, avec et sans coiffe en émail, et en vues mésiale, buccale, distale et linguale. Les facettes d’usure interproximales mésiales et distales sont visibles sur les vues vestibulaire et linguale. ( B ) Une construction d’arbre phylogénétique non racinée, basée sur la forme moyenne de chaque groupe d’hominines en utilisant la méthode de jonction des voisins. NR2 M 2 est proche des Néandertaliens et éloigné d’H. sapiens . ( C ) Canaux radiculaires (cavités pulpaires) dans les vues mésiale, buccale, distale et linguale. Les racines pyramidales et la cavité pulpaire taurodontique se prolongent dans l’apex avant de se ramifier en canaux radiculaires courts. ( D) Les racines dentaires (en marron) et leurs canaux (en rouge) sont présentés en images « verre » de la mandibule en vue latérale, révélant la présence de taurodontisme. Le NR-2 M 2 a une seule racine pyramidale bifurquant au quart apical de la racine ( Fig. 4, C et D ). La grande cavité pulpaire s’étend jusqu’au milieu de la racine et se ramifie en canaux radiculaires courts qui s’étendent jusqu’aux apex, une configuration des racines connue sous le nom de taurodontisme (Fig. 4). Cette racine pyramidale, à cavité pulpaire taurodontique, est fréquente chez les Néandertaliens . Chez l’homme moderne, les deuxièmes molaires inférieures possèdent des racines mésiales et distales séparées avec quelques variations dans les canaux. La racine du NR-2 M 2 ( Fig. 4 et fig. S15) est relativement longue (16,4 mm), tombant vers l’extrémité supérieure de la plage de variation des deux Paléolithique supérieurH. sapiens (11,3 à 16,8 mm) et Néandertaliens (14,3 à 16,5 mm).
Le NR-2 M 2 a une seule racine pyramidale bifurquant au quart apical de la racine ( Fig. 4, C et D ). La grande cavité pulpaire s’étend jusqu’au milieu de la racine et se ramifie en canaux radiculaires courts qui s’étendent jusqu’aux apex, une configuration des racines connue sous le nom de taurodontisme (Fig. 4). Cette racine pyramidale, à cavité pulpaire taurodontique, est fréquente chez les Néandertaliens . Chez l’homme moderne, les deuxièmes molaires inférieures possèdent des racines mésiales et distales séparées avec quelques variations dans les canaux. La racine du NR-2 M 2 ( Fig. 4 et fig. S15) est relativement longue (16,4 mm), tombant vers l’extrémité supérieure de la plage de variation des deux Paléolithique supérieurH. sapiens (11,3 à 16,8 mm) et Néandertaliens (14,3 à 16,5 mm). L’analyse 3D GM pour la forme de la couronne dentinaire (configuration des repères combinant les informations de la jonction émail-dentine ou EDJ, et celles de la jonction cémento-émail ou CEJ : fig. S16 pour le gabarit de mesure, tableau S6 pour les définitions des repères, et fig. S17 pour la parcelle PC1-PC2 et la parcelle PC1-PC3) a montré que le NR-2 M 2 tombe à la marge distante supérieure de la gamme néandertalienne, à proximité des spécimens de Krapina et d’Ehringsdorf G. La variation de forme le long de PC1 (30,6 % de la variance totale) est déterminée par la hauteur relative de la couronne et par l’expansion bucco-linguale de l’EDJ par rapport au contour de la dentine. Comme le M 2 des Néandertaliens et H. sapiens , NR-2 M 2 présente une couronne relativement haute et un EDJ élargi bucco-lingualement. Le long de PC2 (14,7 %), le NR-2 M 2 trace vers la plage la plus extrême de la distribution, à l’opposé des spécimens H. sapiens , Atapuerca SH et African MP. La forme associée est caractérisée par l’expansion de l’aspect distal de la couronne dentinaire, une caractéristique que NR-2 M 2 partage avec certains spécimens néandertaliens (Krapina et El Sidrón) et le MP européen HomoEhringsdorf G (texte complémentaire E). Contrairement au pariétal et à la mandibule, la construction des arbres phylogénétiques non racinés, basée sur les données combinées CEJ-EDJ ( Fig. 4B ), a abouti à une affiliation claire avec les Néandertaliens, tandis que Qesem QC-J15 était associé à Atapuerca SH. Concernant la taille de la couronne, NR2 M 2 est en dehors de la gamme humaine moderne (fig. S18).
L’analyse 3D GM pour la forme de la couronne dentinaire (configuration des repères combinant les informations de la jonction émail-dentine ou EDJ, et celles de la jonction cémento-émail ou CEJ : fig. S16 pour le gabarit de mesure, tableau S6 pour les définitions des repères, et fig. S17 pour la parcelle PC1-PC2 et la parcelle PC1-PC3) a montré que le NR-2 M 2 tombe à la marge distante supérieure de la gamme néandertalienne, à proximité des spécimens de Krapina et d’Ehringsdorf G. La variation de forme le long de PC1 (30,6 % de la variance totale) est déterminée par la hauteur relative de la couronne et par l’expansion bucco-linguale de l’EDJ par rapport au contour de la dentine. Comme le M 2 des Néandertaliens et H. sapiens , NR-2 M 2 présente une couronne relativement haute et un EDJ élargi bucco-lingualement. Le long de PC2 (14,7 %), le NR-2 M 2 trace vers la plage la plus extrême de la distribution, à l’opposé des spécimens H. sapiens , Atapuerca SH et African MP. La forme associée est caractérisée par l’expansion de l’aspect distal de la couronne dentinaire, une caractéristique que NR-2 M 2 partage avec certains spécimens néandertaliens (Krapina et El Sidrón) et le MP européen HomoEhringsdorf G (texte complémentaire E). Contrairement au pariétal et à la mandibule, la construction des arbres phylogénétiques non racinés, basée sur les données combinées CEJ-EDJ ( Fig. 4B ), a abouti à une affiliation claire avec les Néandertaliens, tandis que Qesem QC-J15 était associé à Atapuerca SH. Concernant la taille de la couronne, NR2 M 2 est en dehors de la gamme humaine moderne (fig. S18).
Les preuves cumulatives des trois éléments anatomiques analysés (os pariétal, mandibule et M 2 ) révèlent une combinaison unique de caractéristiques archaïques et néandertaliennes, soutenant l’existence d’une population locale levantine au MP final. Les résultats des analyses discriminantes quadratiques (AQD) (tableau S7) renforcent ce constat, montrant qu’une affiliation des fossiles NR avec des H. sapiens précoces et récents est hautement improbable, mais qu’il est impossible d’établir si les fossiles NR sont plus probables. à classer comme MP Homo , Neandertal ou H. erectus (ce dernier pour le pariétal uniquement). Par conséquent, le tracé de la fonction discriminante (fig. S1) montre que le pariétal NR-1 tombe entre le H. erectus/ Groupe MP Homo africain et MP Homo européen/ Néandertaliens, avec une probabilité similaire d’appartenir à l’un ou l’autre groupe ( H. erectus = 0,41, MP Homo = 0,34, Néandertal = 0,25, sur la base des trois premiers PC).
Les premières caractéristiques de Néandertal dans les fossiles du Levant ont été discernables dans le MP il y a environ 400 ka à la grotte de Qesem , les premiers humains modernes étaient présents au Levant il y a environ 180 ka, et les Néandertaliens sans équivoque n’apparaissent pas dans le Moyen-Orient avant il y a environ 70 000 ans. NR comble une lacune dans ce registre, en affichant une morphologie très hétérogène, mais archaïque. Le pariétal documente une forme plutôt archaïque du casse-tête ; la mandibule est semblable à celle de MP Homo ; la molaire ressemble assez à celle de Néandertal, semblable à Ehringsdorf G.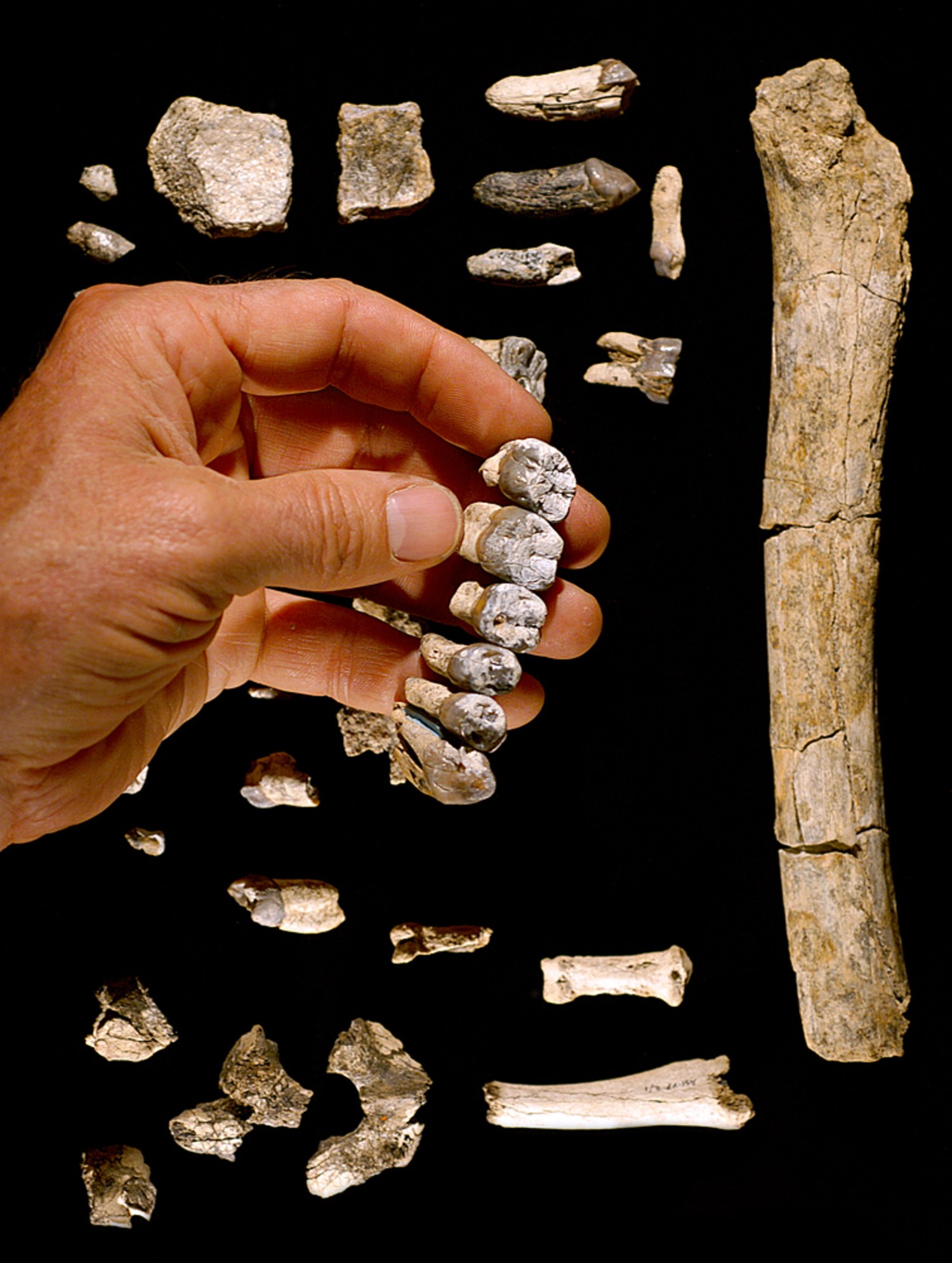 Arsuaga et al . ont préconisé un développement évolutif plus précoce de l’appareil masticateur, par rapport au casse-tête chez les Néandertaliens. De même, les fossiles de Jebel Irhoud d’Afrique du Nord possèdent un neurocrâne plus primitif mais un visage et une dentition plus H. sapiens . Des populations archaïques portant des caractéristiques de type Néandertal étaient également présentes dans une grande partie du continent eurasien pendant le MP, révélées par les découvertes chinoises de Maba, Xujiayao et Xuchang. L’existence de populations asiatiques MP s’écartant nettement du paradigme de H. erectus a été proposée à plusieurs reprises, par exemple pour les dents de Tongzi ou le crâne de Sambungmacan 3 ; ce dernier (avec Ngandong 6 et 7) montre de fortes affinités morphologiques avec les pariétaux NR-1.
Arsuaga et al . ont préconisé un développement évolutif plus précoce de l’appareil masticateur, par rapport au casse-tête chez les Néandertaliens. De même, les fossiles de Jebel Irhoud d’Afrique du Nord possèdent un neurocrâne plus primitif mais un visage et une dentition plus H. sapiens . Des populations archaïques portant des caractéristiques de type Néandertal étaient également présentes dans une grande partie du continent eurasien pendant le MP, révélées par les découvertes chinoises de Maba, Xujiayao et Xuchang. L’existence de populations asiatiques MP s’écartant nettement du paradigme de H. erectus a été proposée à plusieurs reprises, par exemple pour les dents de Tongzi ou le crâne de Sambungmacan 3 ; ce dernier (avec Ngandong 6 et 7) montre de fortes affinités morphologiques avec les pariétaux NR-1.
Les fossiles NR pourraient représenter des exemples survivants tardifs (140 à 120 ka) d’un groupe MP Homo distinctif d’Asie du Sud-Ouest , antérieur aux Néandertaliens levantins d’Amud, Kebara et Ein Qashish (70 à 50 ka). Sur la base de leur morphologie en mosaïque montrant un degré différent de caractéristiques néandertaliennes, d’autres fossiles MP Levantin, dont les affinités taxonomiques ont longtemps été débattues, des sites de la grotte de Qesem ( 19 ), de la grotte de Zuttiyeh ( 30 ) et probablement de la grotte de Tabun , pourraient également être attribués à ce groupe (texte complémentaire F). Adoptant l’approche prudente prônée par Mayr , nous suggérons d’aborder ce paléodème MP levantin comme le « Nesher Ramla Homo.” Sa présence il y a environ 420 à 120 ka dans une zone géographiquement restreinte a peut-être permis des croisements répétés avec des populations humaines modernes telles que les habitants de la grotte de Misliya , une notion également soutenue par leur tradition technologique commune [texte complémentaire F]. Ce scénario est compatible avec la preuve d’un flux génétique précoce (il y a 200 à 400 ka) entre les humains modernes et les Néandertaliens et aide à expliquer l’expression variable des caractéristiques dentaires et squelettiques des fossiles levantins ultérieurs des populations Skhul et Qafzeh. , phénomène constaté par les anthropologues depuis les années 1930. De plus, une étude récente des restes dentaires d’Atapuerca SH et d’Arago a suggéré l’existence de plus d’une lignée Homo dans MP Europe [voir aussi ] et a émis l’hypothèse de la contribution des groupes Homo Levantins porteurs de traits de type Néandertalien à l’Europe. Homo lignées. L’ Homo NR , porteur de traits proches de ceux de Néandertal, pourrait ainsi représenter la population « source » postulée dans le modèle démographique « sources et puits », selon lequel l’Europe occidentale s’est repeuplée par une série de migrations successives.
Ce scénario est compatible avec la preuve d’un flux génétique précoce (il y a 200 à 400 ka) entre les humains modernes et les Néandertaliens et aide à expliquer l’expression variable des caractéristiques dentaires et squelettiques des fossiles levantins ultérieurs des populations Skhul et Qafzeh. , phénomène constaté par les anthropologues depuis les années 1930. De plus, une étude récente des restes dentaires d’Atapuerca SH et d’Arago a suggéré l’existence de plus d’une lignée Homo dans MP Europe [voir aussi ] et a émis l’hypothèse de la contribution des groupes Homo Levantins porteurs de traits de type Néandertalien à l’Europe. Homo lignées. L’ Homo NR , porteur de traits proches de ceux de Néandertal, pourrait ainsi représenter la population « source » postulée dans le modèle démographique « sources et puits », selon lequel l’Europe occidentale s’est repeuplée par une série de migrations successives.
Homo qui ? Une nouvelle espèce humaine mystérieuse a été découverte en Israël Un groupe international d’archéologues a découvert une pièce manquante dans l’histoire de l’évolution humaine.
Un groupe international d’archéologues a découvert une pièce manquante dans l’histoire de l’évolution humaine.
Comme le détaillent les chercheurs Israel Hershkovitz, Yossi Zaidner et leurs collègues dans deux études complémentaires publiées dans Science, cette communauté humaine archaïque a échangé à la fois sa culture et ses gènes avec des groupes d’Homo sapiens voisins pendant plusieurs milliers d’années.
Les nouveaux fossiles
Des morceaux d’un crâne, y compris un pariétal droit (vers l’arrière / le côté du crâne) et une mandibule presque complète (mâchoire) étaient datés de 140 000 à 120 000 ans, l’analyse révélant que la personne à qui il appartenait n’était pas entièrement H sapiens .
Cependant, ils n’étaient pas non plus des Néandertaliens, qui était le seul autre type d’humain à avoir vécu dans la région à l’époque.
Au lieu de cela, cet individu tombe en plein milieu : une population unique d’Homo jamais reconnue auparavant par la science.
Grâce à une comparaison détaillée avec de nombreux autres crânes humains fossiles, les chercheurs ont découvert que l’os pariétal présentait des traits «archaïques» qui sont sensiblement différents des H. sapiens anciens et récents. De plus, l’os est considérablement plus épais que ceux trouvés à la fois chez les Néandertaliens et la plupart début H. sapiens.
La mâchoire présente également des caractéristiques archaïques, mais comprend également des formes couramment observées chez les Néandertaliens.
Les os révèlent ensemble une combinaison unique de caractéristiques archaïques et néandertaliennes, distinctes à la fois des premiers H. sapiens et des Néandertaliens ultérieurs.
Y a-t-il plus de ces personnes?
Les auteurs suggèrent que des fossiles trouvés sur d’autres sites israéliens, dont la célèbre Dame de Tabun, pourraient également faire partie de cette nouvelle population humaine, contrairement à leur identification précédente à Néandertal ou à H. sapiens.
La « Dame de Tabun » (connue des archéologues sous le nom de Tabun C1) a été découverte en 1932 par l’archéologue pionnière Yusra et sa directrice de terrain, Dorothy Garrod.
Abondamment étudié, cet important spécimen nous a beaucoup appris sur l’anatomie et le comportement de Néandertal à une époque où l’on en savait très peu sur nos énigmatiques cousins évolutionnaires.
Si Tabun C1 et d’autres des grottes de Qesem et de Zuttiyeh étaient bien des membres du groupe Nasher Ramel Homo, cette réanalyse expliquerait certaines incohérences dans leur anatomie précédemment relevées par les chercheurs. Le mystérieux Nesher Ramla Homo pourrait même représenter notre plus récent ancêtre commun avec les Néandertaliens. Son mélange de traits soutient la preuve génétique que le flux génétique précoce entre H. sapiens et les Néandertaliens s’est produit il y a entre 400 000 et 200 000 ans. En d’autres termes, ce métissage entre les différentes populations d’Homo était plus courant qu’on ne le pensait auparavant.
Le mystérieux Nesher Ramla Homo pourrait même représenter notre plus récent ancêtre commun avec les Néandertaliens. Son mélange de traits soutient la preuve génétique que le flux génétique précoce entre H. sapiens et les Néandertaliens s’est produit il y a entre 400 000 et 200 000 ans. En d’autres termes, ce métissage entre les différentes populations d’Homo était plus courant qu’on ne le pensait auparavant.
Encore plus déroutant, l’équipe a également trouvé une collection de quelque 6 000 outils en pierre sur le site de Nesher Ramla.
Ces outils ont été fabriqués de la même manière que les groupes H. sapiens contemporains ont fabriqué leur technologie, avec une similitude si forte qu’il semble que les deux populations – Nesher Ramla Homo et H. sapiens – traînaient régulièrement. Il semble qu’ils n’échangeaient pas seulement des gènes, mais aussi des conseils sur la fabrication d’outils.
Et il y a eu le feu !
Le site produisait également des ossements d’animaux capturés, abattus et consommés sur place. Ces découvertes indiquent que Nesher Ramla Homo a chassé une gamme d’espèces, y compris la tortue, la gazelle, l’aurochs, le sanglier et l’autruche.
De plus, ils utilisaient le feu pour cuisiner leurs repas, comme en témoigne la découverte d’un feu de camp du même âge que les fossiles. En effet, les Nesher Ramla Homo ne ramassaient pas seulement du bois pour faire des feux de camp et cuisiner, mais géraient aussi activement leurs feux comme les gens le font aujourd’hui. Alors que les premières indications d’utilisation contrôlée du feu sont beaucoup plus anciennes – il y a peut-être un million d’années – la chose intéressante à propos de ce feu de camp particulier est la preuve que les gens de Nesher Ramla l’ont entretenu avec autant de soin que les H. sapiens et les Néandertaliens contemporains ont fait leurs propres feux.
Alors que les premières indications d’utilisation contrôlée du feu sont beaucoup plus anciennes – il y a peut-être un million d’années – la chose intéressante à propos de ce feu de camp particulier est la preuve que les gens de Nesher Ramla l’ont entretenu avec autant de soin que les H. sapiens et les Néandertaliens contemporains ont fait leurs propres feux.
Le plus impressionnant est que le feu de camp a survécu, intact, en dehors d’un environnement de grotte protégé pendant si longtemps. C’est maintenant le plus ancien feu de camp intact jamais trouvé à l’air libre.
En somme, si nous pensons à l’histoire de l’évolution humaine comme une bibliothèque Ikea qui n’arrive pas tout à fait à se mettre en place, cette découverte revient en fait à trouver l’étagère manquante enfouie au fond de la boîte. Le nouveau Nesher Ramla Homo permet une structure mieux ajustée, même si quelques mystérieuses pièces « supplémentaires » restent à méditer.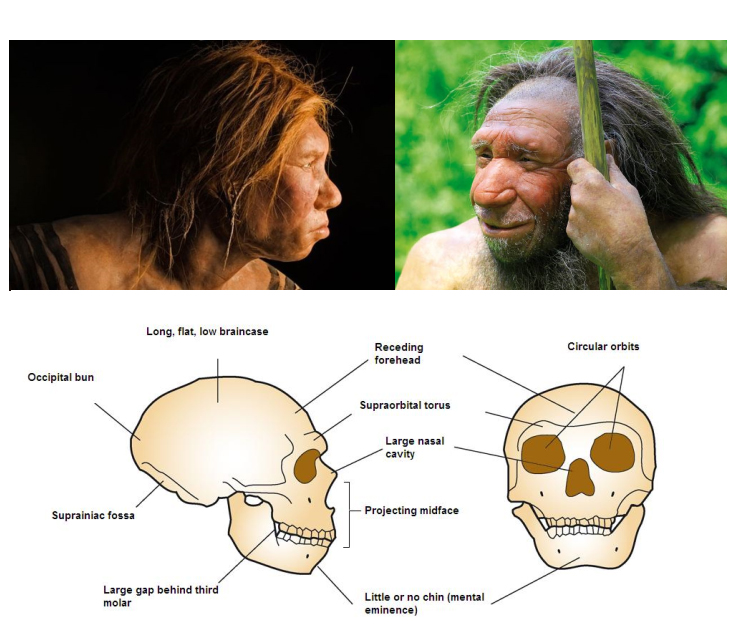 Par exemple, comment exactement les différents groupes Homo ont-ils interagi les uns avec les autres ? Et qu’est-ce que cela signifie pour les changements culturels et biologiques qui se produisaient pour les populations Homo à cette époque ?
Par exemple, comment exactement les différents groupes Homo ont-ils interagi les uns avec les autres ? Et qu’est-ce que cela signifie pour les changements culturels et biologiques qui se produisaient pour les populations Homo à cette époque ?
Un nouveau type d’humain ancien est découvert en Israël
Les archéologues ont mis au jour un nouveau type d’humain préhistorique qui a émergé il y a 400 000 ans et qui se serait probablement croisé avec Homo sapiens. Des fragments de crâne et de mâchoire d’un Homo ‘Nesher Ramla’ ont été trouvés sur un site préhistorique en plein air du même nom dans une cimenterie près de la ville de Ramla, en Israël.
Des fragments de crâne et de mâchoire d’un Homo ‘Nesher Ramla’ ont été trouvés sur un site préhistorique en plein air du même nom dans une cimenterie près de la ville de Ramla, en Israël.
Des chercheurs de l’Université de Tel-Aviv et de l’Université hébraïque de Jérusalem ont daté les restes il y a environ 140 000 à 120 000 ans.
Les grandes dents, la structure du crâne et l’absence de menton de Nesher Ramla Homo rendent sa morphologie nettement différente de celle des humains modernes, a révélé une analyse.
Mais il a des caractéristiques en commun avec les Néandertaliens – en particulier ses dents et sa mâchoire – tandis que son crâne ressemble à celui d’autres spécimens archaïques d’Homo. Ainsi, la découverte pourrait répondre à une grande énigme de l’évolution humaine – comment les Néandertaliens en sont venus à avoir des gènes Homo sapiens bien avant que les deux groupes ne se rencontrent en Europe.
Ainsi, la découverte pourrait répondre à une grande énigme de l’évolution humaine – comment les Néandertaliens en sont venus à avoir des gènes Homo sapiens bien avant que les deux groupes ne se rencontrent en Europe.
Les chercheurs affirment que le peuple Nesher Ramla pourrait être la population « manquante » précédemment supposée qui s’est accouplée avec les humains modernes il y a 200 000 ans.
En fait, ils ont probablement précédé les humains modernes au Levant de 200 000 ans avant de se chevaucher pendant plus de 100 000 ans par la suite.
L’équipe soupçonne que le peuple Nesher Ramla était la source à partir de laquelle de nombreux humains du Pléistocène moyen se sont développés, y compris les Néandertaliens «européens». « La découverte d’un nouveau type d’Homo est d’une grande importance scientifique », a déclaré l’auteur de l’article et anthropologue biologique Israel Hershkovitz de l’université israélienne de Tel Aviv.
« La découverte d’un nouveau type d’Homo est d’une grande importance scientifique », a déclaré l’auteur de l’article et anthropologue biologique Israel Hershkovitz de l’université israélienne de Tel Aviv.
« Cela nous permet de donner un nouveau sens aux fossiles humains découverts précédemment, d’ajouter une autre pièce au puzzle de l’évolution humaine et de comprendre les migrations des humains dans l’ancien monde », a-t-il ajouté.
« Même s’ils vivaient il y a si longtemps, à la fin du Pléistocène moyen (il y a 474 000 à 130 000 ans), les Nesher Ramla peuvent nous raconter une histoire fascinante, révélant beaucoup sur l’évolution et le mode de vie de leurs descendants. »
Les restes ont été retrouvés lors d’une fouille dans la zone minière de la cimenterie de Nesher. En creusant à 26 pieds de profondeur, les chercheurs ont également trouvé de grandes quantités d’os d’animaux – notamment des chevaux, des daims et des aurochs ressemblant à des bœufs – et des outils en pierre.
En creusant à 26 pieds de profondeur, les chercheurs ont également trouvé de grandes quantités d’os d’animaux – notamment des chevaux, des daims et des aurochs ressemblant à des bœufs – et des outils en pierre.
L’analyse de la morphologie osseuse a confirmé qu’ils appartenaient à un nouveau type d’Homo – le premier à être classé sur la base de restes découverts en Israël.
« C’est une découverte extraordinaire. Nous n’avions jamais imaginé qu’aux côtés d’Homo sapiens, l’Homo archaïque parcourait la région si tard dans l’histoire humaine », a déclaré l’auteur de l’article et archéologue Yossi Zaidner de l’Université hébraïque de Jérusalem.
« Les découvertes archéologiques associées aux fossiles humains montrent que « Nesher Ramla Homo » possédait des technologies avancées de production d’outils en pierre et interagissait très probablement avec l’Homo sapiens local. » «Avant ces nouvelles découvertes, la plupart des chercheurs pensaient que l’homme de Néandertal était une « histoire européenne »», a déclaré le professeur Hershkovitz.
«Avant ces nouvelles découvertes, la plupart des chercheurs pensaient que l’homme de Néandertal était une « histoire européenne »», a déclaré le professeur Hershkovitz.
La théorie dominante, a-t-il expliqué, proposait que « de petits groupes de Néandertaliens aient été forcés de migrer vers le sud pour échapper à la propagation des glaciers, certains étant arrivés en Terre d’Israël il y a environ 70 000 ans ».
« Les fossiles de Nesher Ramla nous font remettre en question cette théorie, suggérant que les ancêtres des Néandertaliens européens vivaient au Levant il y a déjà 400 000 ans, migrant à plusieurs reprises vers l’ouest vers l’Europe et vers l’est vers l’Asie. »
« En fait, nos découvertes impliquent que les célèbres Néandertaliens d’Europe occidentale ne sont que les vestiges d’une population beaucoup plus importante qui vivait ici au Levant – et non l’inverse. »
« Les gens pensent en paradigmes. C’est pourquoi des efforts ont été faits pour attribuer ces fossiles à des groupes humains connus comme Homo sapiens, Homo erectus, Homo heidelbergensis ou les Néandertaliens », a déclaré l’auteur de l’article, Rachel Sarig.
« Mais maintenant, nous disons: » Non « . C’est un groupe en soi, avec des traits et des caractéristiques distincts », a ajouté l’anthropologue dentaire de l’Université de Tel Aviv. « À un stade ultérieur, de petits groupes du type Nesher Ramla Homo ont migré vers l’Europe – où ils ont évolué pour devenir les Néandertaliens « classiques » que nous connaissons – et aussi vers l’Asie, où ils sont devenus des populations archaïques avec des caractéristiques de type Néandertalien. »
« À un stade ultérieur, de petits groupes du type Nesher Ramla Homo ont migré vers l’Europe – où ils ont évolué pour devenir les Néandertaliens « classiques » que nous connaissons – et aussi vers l’Asie, où ils sont devenus des populations archaïques avec des caractéristiques de type Néandertalien. »
Le membre de l’équipe Gerhard Weber – un anthropologue évolutionniste de l’Université de Vienne – a accepté, ajoutant qu’il pensait que l’histoire de l’évolution de Néandertal serait racontée différemment maintenant.
« L’Europe n’était pas le refuge exclusif des Néandertaliens d’où ils se diffusaient occasionnellement en Asie occidentale », a-t-il déclaré.
« Nous pensons qu’il y a eu beaucoup plus d’échanges latéraux en Eurasie, et que le Levant est géographiquement un point de départ crucial, ou du moins une tête de pont, pour ce processus. »
https://en.kataeb.org/articles/arts-culture-2021-06-25-a-new-type-of-ancient-human-is-discovered-in